Rôle de la régulation génique dans l adaptation : approche par analyse comparative du transcriptome de drosophile
|
|
|
- Ella Harmon
- 5 years ago
- Views:
Transcription



1 Rôle de la régulation génique dans l adaptation : approche par analyse comparative du transcriptome de drosophile François Wurmser To cite this version: François Wurmser. Rôle de la régulation génique dans l adaptation : approche par analyse comparative du transcriptome de drosophile. Sciences agricoles. Université Paris Sud - Paris XI, Français. <NNT : 2011PA112260>. <tel > HAL Id: tel Submitted on 3 Jan 2012 HAL is a multi-disciplinary open access archive for the deposit and dissemination of scientific research documents, whether they are published or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.


2 UNIVERSITE PARIS-SUD 11 ÉCOLE DOCTORALE Gènes, Génomes, Cellules Biologie évolutive THÈSE DE DOCTORAT Pour obtenir le grade de Docteur en Sciences de l'université Paris 11 par François WURMSER Rôle de la régulation génique dans l'adaptation : approche par analyse comparative du transcriptome de drosophile Le 25 novembre 2011 Composition du jury : Président du jury : Rapporteurs : Examinatrice : Directrice de thèse : Co-directrice de thèse : Dominique De Vienne Frédéric Fleury Bruno Lemaître Carole Smadja Dominique Joly Catherine Montchamp-Moreau
3
4
5
6 r ts rr à r rt s à é r r s rs r r r s r s r2 t ès t t s 1 r rt rs ré ér r2 t r îtr s ss q t r r r q 2 r r s rté s s ts t s s r rr é s r ss èr tés s r tt t r s t t t à s tt t ès s t s à s rt t s é t à t s à r t r r r s t t r t r t rs à t é tér ss t 2 sé t r t s ss tt t ès s r s r t ré té r r r r 3 r s t s t t s t t s s s ét s t s s r t q st t st q t s ût r s ss t s r tt s r r r r P r q s s s r r st t r q t é r t t t r t t t s rt t t rs s r t r ê à s s t st q ê q s à étr r r r r s st t 1 q s s ré r r ré ss à r é t q ét s t q s s r t t q q s st t st q s s ré t t rr sé st t s r s q s à sûr s s rt t r st r r r é tr s és st é P r q é tr r s é t t ât t t t ré t s s â ê s q q r r r r t tr ès t t r r é t ç s t t 3 à tr t ré éré r r r s t r r q s s s r s r s à P r r
7 r r t str à s r t r q râ r q s t q r r r r t s s t és r s q s t ssés t tt t ès r q r t s s s ê èr s êt s sûrs q st r à t é ïs s rs t ès r é t é s rt t r s s t t s tt s ss é s s r s ss s t t r ît s ô s t st s t r ë é r t r ç s 2 r r P rr t r ss2 q t r q é s r r s s str r s s s r tr é 2 t t é r r é 2 t t t ré t s s st r s s s à îtr t t q 2 rr é é r t t é ré 2 q s rr t s s à t t èr P rr s s q s r s s é r ç s r ç s s s r r t s r t s ût r t s 2 rs q t r t 2 r r r r s st 2 st r q r s r ér q ë ï ï s s rs t t ét q P tr tr r 1 à s é è r r st r ç s s r rt2 r r r t é à r t str 2 t r s t r r t s t ât t ât t r r s sé ré s s s s t rs é st r é t r ôté s t s rt èr t q é q é s s r r t t r t r séq ç t é t r t r s s é tr r P t r t r è s é s s t rs té P r s s P r tér q t r r ü s r r r P rr 2 q t t t r q ss r tt t ès s s r t r r r r s s s r r é s r r t
8 é è P rt r r r P rr r 2 r t ts ss s r t st s s r s s q és é é ê t s r ê t s t 2 r r r t t s r ts é s t rs à r r r q r à rs r s rô s t q s s à r 1 s r s r ts t rs t t s r s s r rèr é ré ré s s s ë é 2 t P t t ss é r s t s s t s rèr 2 r
9 s t èr s tr t s è tr é t s t r r t r é t t r tér st q s s t ré32 t q s t st32 t q 1 r ss t é t r 1 r ss s s t s 1 r ss r t t 1 r ss t é é ts tr s s s t q s r t s 1 r ss éré té t 1 r ss r 1 r ss tr t 1 s t s s è s tr è r s s s rs s é q tr t r t s t s s ét s 1 r ss 3 s s tér t ét s r s s s s s t rs 2 r s r s à é s r s s t t s é t s 1tr t P s à t 2 r t r s t 2s st t st q étér s s
10 r s s r t r é s s r ss t rr r t s è s 2s tr s é q 3 s s r séq ç é ér t t s é t s s s 1tr t Pré r t r r t séq ç rt r s séq s r s rt é é ts tr s s s r P 2s st t st q t s è s és t ts t s ss r s s tr t r s é q s t 2 r s s s à s ér t à t s t s étér s é q s r s s 2 r s tr t s r t s t rt r t 1 r ss 3 s 2 r s è s ér t t 1 r és tr 2 r s t r ts s t s è s rt r és s r r rés t t r s ô ré t s étér s s r s s è s t s ér s 2s t 2t r P r â t sé t r ét 1 t 3 s é t r r s t s 3 s s ér t 1 r ss tr t s s s r tr t ér s é s à st r s t s t s t s s t q s séq ç r ét r 1 r ss 3 s t s s s tr té rt r rq t
11 r s t s t t à r t ô s t t tr s ér s s s t t r 2t r s P 2 st r é t r t è s q ré 1 str ss ts r ss r s séq s r s à 1é q t té 1é q à t s s r r s s s ê t 1é q à ût r r r t 1é q à r è s és s ss é ér ér t 1 r ss tr t s s s ér t r s tr s 1 ét s 1 r ss t t t s 2t r s P rt t é à t t ss t t rô s 2t r s è s ts t à 1 r ss é r r t 1 s 2t r s tr t t t s é é ts tr s s s t t q é é ts t è s és à t t s r t t 1 r ss t é t tr q s ét 1 r ss Pr s é ér 1 s s séq ç t é t Pr 1 s ts t rts tt t ès P rs t s r t q s q s r s r 1 é ts t rs tr s r t r tr s r t t rs tr s r t ét 1 t t t s t r s st 2 t ès é s t
12 s r s ér t s r s t r r ré rt t s é s s s t t str t s s t st é é té r str t s s t st s r s é s s é s ét t s rt r P tr 1 P r ètr s r s rs t 2 1 r ss str t s s r r s 1 r ss t s str t s ér s r è s ér t t 1 r és r r rt à è s é t r s str t s r rt d/a r s q tr r s ts 2 r s ét s è s ér t t 1 r és tr s t s s s r t2 s t t rès r2 t P r t tr s r ts rt r és str t t 1 r r s è s tés rt s tr st r t s s r s è s ss és é t r t str t t 1 r s è s rt s rès rr t s t r s q s è s s r 1 r és à t r r r rt à 2 tt rts 1 r ss t r s r 2 tt r s è s s 1 r és à t r é t t s è s 2 s 3 st r t s s t t s t r s s r r r rt à 1é q t rès t tér q t 1 r ss r s t 2t r s q és s s ét 1 t s
13 st s t 1 P 1 ér s s r s tr q tr t s s s s t q tr t s 2 r s t t st q s séq ç t rt r r s q tr é t s è s s t t s r 1 r és 3 s 2 r s r r rt 1 r ts t s t r s q r è s ér t t 1 r és tr s t s s r s ér t s t s s s r s t 2 s r r rés tés s s è s s r 1 r és 3 s s r r rt à s q q s t t s s è s ér t t 1 r és tr t s s s P s t r t s r t s r s r s s r tr t s r è s ér t t 1 r és tr q r s 1 à 1 s t s r s t s r r rés tés r s è s s r 1 r és à t r r r rt à 2 tt r s t è s s r r rés tés r s è s s r 1 r és s r r r rt à 1é q r s t s è s s r r rés tés s st è s s r 1 r és s r 1é q r r rt à és é s t s ss é s 1 q tr s 2t r s P 3 st r
14 tr t t t s ù s t q s s è s r r t té t t s r s s P s tt tt t s st tr s éré rs s séq s é s ts q ét tré 1 té s t r t s t rs r ss t t s é s rô s r 1 r 1 s ts s r s s s t s r t t s ss s r é t térêt r é r2 t r rô séq s s s t s q s st t rs rés t s tt r èr é t 1 r ss é q s s t é t s r s s r s r ss t rô 1 r ss s è s t ré sé t s t t s ér t s t s s s è s tt tr t à rt r t s è s è r à à tr r rt 1 r ss é q t q été ré é é r t r té s t s t r s s è tr é t t s è st à s s r é t q s é t s t s q s t r ét r t s s è s r s 2 é é r t r q s s r ss s q s t à r sé r t ér t s t s s é t ré s s r t s q s tr t t tr s é t r t s t s r s t s ré t ét t s é ér t r r t t t é s2stè s tt q st st é t r tt t ès r q s s r é à ét r s t s ér t s 1 r ss r s r t t t s t é t t à sé r t s t s s s t r r à s é t s r t rt st st q s r t s 1 r ss tr s t 1 s t s è rt rt s t à s 1 r s s é t s
15 s è rs s â s t à térêt rs r s s è s q st r t r rt t r r s ê ré s s è q st é r t s è st s t r t r sq à P t r r èr r IV ème s è t é t s t2 s ér ts t st t r té à s 2 r r t é t r séq t s è r é rs q st tr t q t s s 2 r t ss t r t t t s è st str à é t t été r é é t t r rt tr st r sq XVIIème s è s èr q 2 t t s è s à r t q à ré t tt t st ss t t à rés r s s rsq é r à ss r s s è s r t à r st t r s r s réés 1 r s s s s r s r tèr s r q s t t t q r s 1 s r s r t s r q s r é t s ss t s s êtr s ts t s s s t 1t 1 st s s t 1t é t st r è t s 1 r r t t tt r XVIIIème t s r rs é t st s r r r r r îtr é t sé s r t r é té s êtr s ts st t t s è s t s t s s s t s r r s s ts rt s st é t q s è s r t r r st 2r é é s s r r s t 2 r t t 2 t r r t r t s r s t r t r s r s 2st t s t r s s s 2r tt é t t s s êtr s ts s r r s s ts t r rt s s t ê s è tr r s t s ê s è t s s êtr s ts q s t s és t r r t t r r s s ts rt s 1 st t rs s é t s s è t s s s t t t s sé s s r és tr r t s t t r r s s rr s t à ér t s r s q r r st 2r 2r é q r r t tt t 2 é ét q s r t r é t é q s s r s t r t s s è s r t tt é t st t t ê s è t s s êtr s ts rés t t té é q t ê é q
16 ê s é s2stè t 2 é ét q é t é ér t ré r ss ss r st t r s r s st ts tt s r t êtr sé s r s r tèr s r q s é r s r ê s 2 r s r s 1 P s rs t rs s s t ss 2és à s é t s s s s t s è P r 1 t r é é r s s s r t 2 r t t 2 t r r t r t s t s t r t r s t t r s t t t t t t r r r t t t2 r s ss t t r s t t2 r st t ss t r s r r2 t t 2 t s r s t r 2 tt é t r t s é s é r s é q s t 2 é ét q s rt é s t r t t r rt té rt s s s té é t q s s t s rs à s t s t r r t r é t t r tér st q s é t q s è è t r t à t s t r r t r t s t t ré ér à é s s è s st à r à s 1 è s tr r s t à é t é t s r s s t r réq s é q s t t s t tt t s r rt r s s tt s q r t s 1 q r r t s t r r t r t êtr ré32 t q st32 t q st à r t r t t rès é t tt t r t tt t ès s s 1 é 1 r ss s è s s r stér té té 2 r s t st32 t q s t ré32 t q ré32 t q s t tés t êtr é q s st à r é s à t t r 1 t s ôt s ér t s r s s é s 1 t rs s t rs ttr t té s s r rt s t s r s s t s r à s é s t r s s ér s rt té r s é é q q s r s q q s s s ér r r t é é q q s s r ê t q s r r t
17 ç s P r r r s t t r 1 r é r s t s q s s ré s s s r q s s r r t s t s s t s s 1 s ré t s t r r t r rt s t t s t ré32 t q t ss êtr û à s r è s r ss s rt r s s 1 s rss r t tt t êtr é t s s 1 r ss q s s t s s r t s ts r t t s q s s ts r t rt q é s è 2 r s à tér r ê r s è s r s s è s é st r s s t s â q r t t r tr s è q â q r t s t r rs q s s t st r t t 1 â q r t t r tr s è q â q r t s s ér t rô rt r tèr s s t s r t s t tés é q s é s à t t ss r rô r s s t ré32 t q r r s t t tré q s ér s t 3 é3 r r q t s tés é q s rs t s t s t tré r é s t q t r t r rt r s s r èr s ét s s é t s t ré32 t q t êtr t t é à s r è s r ss tr 2t t s r t 3 ï 3 s 2 P s t st32 t q s t t ss s s t r st 32 t q st à r q 2 r t 32 t s s 2 r s és r s t s r t s t s s t stér s r s st 1 r s ét s s r s t r r t r t s é ér t r s ts s t ss s t r t t s tr s è s r s s è s s t r r t r t t é r ts t é ér t été 3 r s s é è r st è 2ss s t tr r ss ré s t r t rs s s ré stér té P r 3 t s 2 r s s t é ér t è st q s s s 1 s 1 s st stér s r s 1 étér èt q ér t s t é r s t été é s r 1 q r rè t é r é à rés t r t s é étèr s ré ss s s r r s s s2stè
18 é t s r s è s t t 1 r ss st sé rs s 1 r r rt 1 è s s s s s 1 s t st rs t tr st s rq é r s è s t r t s s t s â s s r ss s t 1 q r à s rè r s 1 t â st ét q rés t t é t é t s r s è s â s t tr r tér st q 2s t t s 2 r s st t r r s à ss s rs t é r s 1 s s 1 q t tt s r t stér té ét té s t s t s à s t tés tr r s t s t s s rr t r st ér t q rt s s és à t té q t s r r r t s r rè 2 t ès é à s st rt rs s 1 r t r2 t q s été r sé t rr t r s êtr r é Pr s r s s rt r t s s t s é 32 t 3 2 r 2 t rr s rt r t s t t r s é t s r t ès s t s tr ét s s 1 r 1 r ss 3 s 2 r s r s rs s è s r t s tt 1 ér r t s r ttr ré é r s è s q és s s t tés 3 s 2 r s stér té s â s 1 r ss t é t q st tr tt t ès st rô 1 r ss é q s s r ss s ér t s t s t s s è s r t s t r r t r s ss t t t s r ss é q très t t t t s t t s r sé s t êtr sté s r s s ré t t t r t rt r 1 ré r s t t s r r rt 1 t t s séq t Pr t r 1 s tr s r t s s 3 ér ts s t s Pr t s t rt ré t é q t t ré t s ré t r s é é ts 2s q t és è s ré t tés é t s s 1 s é trés t t ré t é q s t rt t r r s t tés à r s t t r r
19 t Pr t t à é r t t t t 1 r ss r t t st rt t t r rés t 1 r r rt t r ré s s r ss s t t Pr t r s r t r r s t t s s s r t s ts r tt t é t st tr térêt r é t q t r t s ts séq s t s t s é t s 1 r ss s t ts s ré t tr t t s st tr t s r t r té q s tér ss é t à r t t t t r r ttr t é t s r q s rs t t s t s P r q s t t s s t rt t s t é t t q s r s t r t é t2 q éré t r st r q t s é t st s t r r é s rt 1 r t t s s t t s t r s r s è s é t rt t tr s t t été s s é t r rô s r t t s s s s ts é t s t r st rt t r q s t t s 2 t s ts tr 1trê s s t r t é é s r ss s é t q r r ss t t à s s t t q r t t s st q r r t s r é st ss ss q t t s r t r é t2 étr r r tt ré é t r té é ét q r2 t q st à r s s séq s r é t2 s 3 r s t r s t r t sé t t r t s r s ér t s t t s st rô ér s é t 1 r ss s s tér ss r s à s q st s s tt t ès s t 1t r r t t s tér ss t s t é t s ts 1 r ss é q s t 1 ts ts r t t à r t P t t t s é s q séq t t é t2 q s ts ré t t t êtr r t2r rt s t t s s s séq s ré tr s r r rt 1 t t s t s t s s sé st t t rt t s t r ttér t r r 2 tt t ss t s t t s s s séq s ré tr s s t s q t t t ér t s s t t s s s séq s t s s r ts r tt t é r s t r t 1 t r s r r té q s t t s s t t s r séq ré tr 2s q t é è t t s é t2 s st tr sèq t s r sé t t s t s r s t t s ré tr s r 2
20 r r r t s s r r tèr 2 q t t t t ré é tr s r t é t st t r tr s r ts r t t t s r q str t r q st r t r s é q r tr réq t s r ts str t r t s r t s st é ér t s r Pr t s r tér st q s 2 q s r tt t é t s t t s t r r s ré s à s r ts r s s r t sé t s s r s t t s ré tr s s s r s rs st ts r s ét s t tré q ré t st s t s é q q è tt t t tt t q s t t s t s t t s t r s à sé t r étér 32 t rés t é à é t2 ér t ré t s t s é té t ss r q r s s t s t q t t t q t s s s t ss s ù è s 1 r r 1 r ss s s t s r s r s tr t tr é ss té t s r r s ét s tré s s r s r s s ré és s t r s rs é s s t r rés t t r rés t t t s s s r s t r tr t r t é r s ér s ré s tr t s t t r r s ét s s r s r s s r t s r t s ss s t r t t r s t tt tr t s tt é ss té rt t été s r tr s ét s t r s s r st r é ér t 1 s s rt r t r t t r t t t sé s é s s ss t ér é r té s t q 3 é s é t r t r s q s s 1 é s r t té à î r t tt s è st é q rt2 t t t sé é s s r 3 ré t é s é st r t t t s t s é r r rés t r s è r t s r r t r tr t t rt r tr s è s é s té s à s t ré s t ét t é é é ér t s r t r s s t s tr t t é t s s s str ss ré t é à r t str ss q 2s q t s r à q t s é s s t r r rés t t s s t s t s r t
21 ê èr r s s è s r é t êtr t é s rs t s s s è s ét é s t t s s s rés t t str t r t s t ss s é ss té s r r r tr t r t r q r t r s s r s st s r r t à tér r s r s s r t t tré très rt r t tr s t s ér s tr s è s r t s t r r s r t s tr t 1 s t s s è s s rs ét s t r r é à é r s r t s 1 r ss à tér r t s t s r t r t t r t t r r t Pr t r t s t t t 2 t r 2 t t t r r t 2 t s t rsq r 1 r s st tr t t r q s r ss ss 3 tr é s rt t s s t s t r t tt r t r t t t r r r s t é t très r s r s t s ét s é t r t t r t r é tr t 3 ér ts r s s r r s s r s ss t t s s ét s ré é t s 2 r s t r st r r é t é ér t ss s è s s t t r s tr s é s s r t s s rt s t t été tré s r r r t t t r r t s ét s t sé s t t st t st q s s t 1trê t rs s t st r r s ét s q t r t r st q s s r s r t s t r s t r s s s 1 t t t 1trê t rt t rt2 t r t P rs t r t q st s té 1 s s t ss s r r t rs t 1 r ss é é t très r t s t ss s 1 és Pr t r t t 2 t r 2 t tr t r s t s str ss t é t rô rt q s t t è s rés st s2stè t r è s és str ss Pr t r t r t s t t 1 r ss r t t P rq ét r 1 r ss s è s rt s t s ér r 1 r ss r r é t2 s s s é t2 q st séq q s r t é t2
22 q st tér ss t st rs à q t s é s 1 r ss s t s r rés t t s ét t t tr s t r s q rré t 2 t tr q t té t q t té r té s s ét s r 1 r ss à é é t r s t t 1 é q tr s ts t r s r r 2 s r s ré és t s s t s t t sé s s rt t s s è s r s à t r s r r r s ér s 1 r ss tr s s ts t t r s è s t rés 1 ré t q és s t tré s r s è s q és s s2 t ès s s és é r t s r té s tr s rt s s ét q s t é t2 r ss s ér s s s é r t tr tr tr t t é r t s r té s t t r t s r q t té r té s r t s r s è s q s t s 1 r t t ré é és r 2s tr s r t q tt q st été s r r 1 é ts s t r t t rs 3 r r t t tré rré t r t t tr q t té t q t té r té s r s t r sé tr s 2 t ès s r 1 q r tt s r t rt ré t s t st tr s r t t tr t 2 rt r s t ss é r t t s2 t ès s r té s t s rt r s s r té s ss s s r tr s r é r s t 2 rr r r t s t s s s r s s s q t tés t r té s q r t s r rré t P ss t s r r r t t é s 2 t ès q s è s s s r s 1 r ss r t tr r r rré t q t té r té s 2 t st é r s ré t é q st t s t t é t q s ré r t t r st à r r té t st t t q s t st té tr t rré t r s tr s r ts s s r s 1 r ss t r s s r s P r s r rs ré t s s t r té q r t t s ét s rés s t q s q t t s s r té s s t t é ér t à r té s r s s t s r st à é r r à q t s s r t s t s s r s s t rt t s r s r té t t é é
23 t q q t t r té q sé s r s t q s s tr étr ss P s t Pr t r ss t t q é à 1 s tr t s r t s s ré t s tr s r t t tr t s t s tré q 3 tér s r r sq t é ré t s r té s tr s r t t q r t à s 3 r s t é rt st r t û à ré t tr s r t r ér 3 tér t s s r t s t é s t t r st t 1 2s s ér t s 1 r ss sq r é t s t rs s è s r s q s r èr r s st à r rès r t s è s r sq s rré t t s r té s st st ss r q r s s r s s q s s2stè ré t ét t s r r s s2stè ér s 1 r ss t é é ts tr s s s s é é ts tr s s s s t 1 ts ts r rt t è 1 r ss s t t tré rt rré t tr r 1 r ss t rés é é ts tr s s s s s rt s t s r s è s t t é t 1 é s ér s 1 r ss 1 s rt s é é t tr s s 2 tr rs q tr s ts t r s r 1 r ss s é é ts tr s s s r st très st à tér r é r s s 1 s é é t q st s ér s 1 r ss s é é ts tr s s s t ré é r t t 1 r ss é st à r r s t 1 r ss t té s r s é é ts ss s s r r t é t2 q 1 r ss s réq t P rt s s r t s é éré s r s é é ts s t s s t é étèr s r r 1 2 s s t t s 1 r ss r sé s rr t r s êtr t s s r ôt t t s é é ts tr s s s r r 1 r ss st é r t s ssè t rs r r s r t rs t é é ts ré t rs s t ss s sér r s s ré s ré tr s é à rés t s r s r s r t s é ss
24 t q s r t s 1 r ss 2s st t st q r t ré é r s ér s 1 r ss st t st q t s t s s q st rs s t s q s t rt s ér s s r é s q q s t r r r t t êtr t s r r sé t t ss t s r é t2 é ér t 1 r ss tr t r ér t st t êtr ét r é q r s r ss s st st q s ér é ét q t t étr t rs r t rs r t t t r rt t s t ê rt rs à ss s ét t st é s r 2s s t é r s ér s s s s P rt t q t êtr s t q ér 1 r ss r 1 s q st s é t s t r t s ré s q 2s t êtr té à q s st t rs s t r rét r s t q r ér s rt t s t r t é r t éré té t 1 r ss s s s té à q t 1 r ss st r s r 1 é t2 s é r s r t q séq t t r q 1 r ss s t rt té r t é t t t t t q s ts s t tr s ss s é ér t à tr r 1 tr 1 t ét é 3 s 2 r s tr t r s é q s s tr s 1 r ss r r rt 1 r ts s t r r r t t té 1 r ss rr s t à 2 1 r ss s r ts t té 1 r ss s r s 1 r ts t étér s s 1 r ss s ér r ér r à s 1 r ts s rés t ts s ét s s t r t tr stés tr t s rt s s t té rt t té s s t r rt2 t t t ê r r s 2 r s s s r t s tt r èr ét s rt t s t tts t t rr r 1 r ss tr t 1 s t s s è s r s s ét s t rté s r s ér s tr t s t s è s s t s s ét s s t rs t ré s r t t t s
25 t s s t t tt r t r t à st t rs té t str t r s t s s t s r t rst 2 t ss t r r r tr t s é t s é s à r s s è s t t t ê t sé 1 r ss rq r s é t s t ré 1 r ss 3 1 s è s r 1 très t ér é s é r t s t s 1 s r s è s t ré é é ss 3 ér t 1 r ss tr s 1 s è s é tr t s t té 1 r ss r s r r s s è s rs s é t tér r ê t s r t s 1 r ss t êtr rt t s st 1trê t r t s ét s t s t 1 s t s ts 1 r ss s t r r és t t q rq r 2 é ét q t s r s ss t st r t été t sé s r r r r s é s s r tr s 1 r ss r s 1 r ss st tr é t à s r t t s 1 t s é s r r é s r t2 r s t s é ér t s s t rs s t rt s t rs t t s éré q r 1 r ss s s 2 t ès é t tr rr t êtr rq r 2 é t t r èr ét q ré sé r s 1 r ss s r s t s t r s rté s r 1 s è s r s ss s té é sté s r s s t s t rs t 1 é r s à tr s r t q s 1 t s t r t s s q q s s è s r r s s t ré é r è s ér t t 1 r és tr s ê t t è s ér t t 1 r és tr t s s è s q st s s r rt r tr r r rt à r t r s s s rr s s q ét 2s r t t r rét r s rés t ts r r t t ét rés té r s t tré str t r s t s 1 r ss s t ré é é s tr s 1 r ss r s r r s t t ér t r t s s r ts t s t tr t r t s tt r s à t t à r t t t t ér t r tt ét st r èr à r tré t t à r t 1 r ss èr s r q t t s séq s sq s s 1 t s t r t s r st t s r s tr s q r s t tr s r é t é t êtr û à s r t
26 t 1 r ss r r s é è s é é ét q s s s t sés t été ré és s t r t êtr é és r t r t r s 1 s t s tré q s r ss s é é ét q s r tt t tr s ss ré t é q rès st t r r t 2 r s à tr rs s é ér t s é è r st t s t r é tt ét s t st t q s t s s q r rr t r s t sts t s q s t t r ç s r t r è s ér t t 1 r és à 1 q t r s tr s ét s t t r t st é s s t t r r s é r s r t t 1 rr r tt t s s t t r 2s t è s r rt t 1 s t s r t s r s rés t ts P r 1 s t t t sé s s rt t è s t t s t t sts st t st q s ré sés 907 0,01 = 9,07 s t r è s 1 s t s r st r st ss 3 t s s t 2s r s ét t s è s q q è s s t s tt st q r s r t é t 2 1 s t s r s t è s ér t t 1 r és tr t s q r rés t r t rt t 1 s t s ét s st rt èr t tré s r r r ér s 1 r ss s s t s t r s rst 2 t s t rs t r â é s s s é s t r t s r t r r r é t s r t s à sé t r r t t r s t 1 é s rs t s à 1 ts s t s s t rs t s s r é s r s é s à t t à s ér s rté t r r té s 1 s q r tr r t t r rt à st sé r t s t s tt s s è s t r st t s ét s 2 t 1 é s r s 1 r ss tr t 1 s t tr r rt é t t t 1 r ss r é r t r 1 r s sé t é à t t à t sé t s 1 s rt s ér s té s tr s s 1 s t rt2 t t r t t r rt r 1 r ss s ss êtr tr P s s t 1 s s t é és 2 é ét q t s r té q s ér s 1 r ss s r é s s t tr s s r r
27 tr è r s s s rs s é q 1 s s st t 1 1 r s ét s é t s à s r r ss s t s s s r r s é ér r r 1 té tr s tr s s è s 1 s s s t r t q t r s 2 r r t r s r tér st q s 1trê t r é s é s s è s s s été é r t r r èr s t rt t r t s q r été t st r tt s è rt t s s r st r t tr s s è s q tr t r rs té é q rt t t r s ré rt t s ér s s t très stré s r s 1 s è s q t été t s ét s s s t s s s st ré t s t q q s 3 s à q q s t s é s s t 3 r s è rr t êtr s t é tr s r t r q st 2 3 s r st t t 2 t ès 1 s t ö t ött r r s st é q s î s 2 s s t t r t s s st é ér st s r r t t s rr t s r s r ss r s r é s rs q s st s é st s rr t t s r r t q s r s r tr t t 1 q r t t s s tr s r s s t s 1 s è s s s t sé ré s st r 2 à s é s 2 t t s t tt r s r èr s st t s t t sé r t s s s t r t 2 s 2 t t s t s s é s s ré t s t t à r s r tt t q s t sé r t s s t s s 1 s st 1trê t ré t à é é t s r s s r s è st é t séq t s s t s r t r r tt t s st s è s é sé s r t r tr s é s ér t s s è s s r tt t t tré q s s ét t 2 s r r s tr s s è s rés t t s rt té t ès r èr r t s t s s s rt té à s t rs ê 1 s t s t s s t r r s rt té r2 r 3 s s t t 1 té st r t à t ïq sé rété r q t té r t tt é rt èr s s té s ét s r r r t s st
28 s r ssé r tt t t s 2s s st té 1 t 1 s t rq s s t à tt t s ét t s r r ss r ss q tr s r s s s st ét t ré t r r s t r t s ré à s s t r t s t t st tt ttr t r r tr s q s té à t ér r s t 1 s tt r èr q t é à s t s r tt r ss r t t éré é q s q s é r tr t r t s t s str t r t é r q s s été t s s r é t s ét s sé s s r s 32 s 22t t r2 t s r s r tèr s é t2 q s 2 t t t à str t r t t s ét s sé s s r 2 r s é r t q r r t tt t tré tt ér t tr s t s r s t r s r q t ér t tr s r s r r q st rr s t à 3 str t r t s t s 2 tt s r 2 t 3 r r q s t r r q tr r rés té r t r s r s s t sé rés é r t r q t s t s r t s t r s t t t q ré t r rs té é r t s t s r é s t é rt t é t s r tér st q s t s é s r r t à rt r 3 str s s r t s s t r é s r s ét s r s t t s q tr t r té ré t s s r s t t s és à r t à s 2 s sé t s ss és à s rt r q s è ö t ött r r s t rs t tré r s t q t t r t s êtr s r s s s tr s s r és t q t r t été ss é à s sé t s s r s s r s t t s s r és t t tt ét à t t t t s 1 t s t s t tr t sr ë ö t ött r r t tré str t r t tr t s r s t r s s é t à tér r r q t s s r s t s r s s t é t tré q s t s r q st t s s t s t s ér é s s rés t ts t été é t r és r ét s r s séq s t s r2 t
29 s s tr 1 tr str t r t tt s t s 3 s s r q t sé r r s t s tr s r s r 1 3 r à st r q st t r q s s 1 r rs ét t és rs ssé t s è t t s t s ér é s q r tr s r s tr s t ts r é r q s s st té à r s 2 s sq à ré t s t tt s è st t é s t r t s s 2s s s s t tré é èr ér t q s s 1 t s r t s ét s 1 r ss 3 s s q s ét s s s t tér ssé s à 1 r ss s tt s è 1 tr s t 1 é rré t tr 2 r s séq t 2 r s 1 r ss tr s s 1 é s séq é s s s 2 t 3 t s t rs t r r é s rré t s tr ér t s rt s s è s tr s t r tr s 1 s t 1 r ss tt r st té r rt r séq ç s s q à 1 r s è s r sq s séq st s s r ss 3 é s s t t tr r q s è s s r s 1 r ss t t rs s r 2 r s séq t t t t s t é t s r é r r t 1 r ss s è s és r s r r rt 1 t s s s t s r é s r r rés t t s è s sés rs s â s 1 r s s s t t s rt t 3 s â s q 3 s s r s s r s 1 r ss t s s s r t s s t ér t s s tr s é ér 1 1 r ss é à t s 1 rt2 t t rt2 t té s ét s 2 t 3 t rés s s rré t tr 2 r s t r té 1 r ss tt rré t rr t s r q sé t s 1 r r è t s r 1 r ss è s r s séq ré tr t s r s séq t s è s s s tr ts 1 r ss s r t 2 r s séq t 1 q é t r 1 r ss ç tr s r t t s r s é r s s tr t s sé t s q r séq s rés t ts s t é t rt 1tré té s ré t é q 2 t 3 t tr s ét s t ré sé s r s s tr s s t tr s r s s 3
30 t t tré é t t t 1 r ss tr s s t st r è s ér t t 1 r és t t t r r è s és à r r t r r s è s és à r r t st tt s s 2 t ès q s é t s s rt s r ss s é s à sé t s 1 ét t r r r t r q s rt rs à r t à s tr s ê s 1 s ss s t t st s tr s 1 s 1 s rss r t r r s é t2 s és à r r t sé t s 1 r q rs s s è s q és s s r ss s s s r t s s t t t é s à é t è s à 1 r ss sé rs s 1 é à é q é s t r t s t tré s r s s ré t è s és à s é s t tr s s t s t s 2s r s s t tt ét s rt rés t t r t r r rs t2 s st tér ssé à 1 r ss 3 s r s s s s r st r t s s t s t s r s st t st q s s t s s rr t r s t sts t s r s rés t ts s r s 2 r s t s s è s s t t s 2 t ès s r t s è s sés rs s 1 t t s â s t r r t t s s s s r s tr s 1 r ss ss q é t s r è s és s 1 è s s r r s tr ts r sé t r é s 2 r s s r r rré t tr é t s t 1 r ss t 1 r ss s s s s r t t r tr t s ss s t r s été ét é t st s s ts r é s tt t ès tr ét s st s é s t 1t s t s t r s 3 r s rt r ss r r rs té 1 r ss à tér r s s è s P r t s t s ét s t é t s st sé s s r ré r s à q tr t s s s tr s r s t r ç s s t rs 2 r s t r s é q s q t ét t r rés té r q tr é s P s s s r r s t 3 str rs s ér é s t q s séq ç t é t q r tt t 1 t rt r tr s r t t t s r ss t s ét s sé s s r 2 r t s tt s rt q t ét t r rés té r s é ts tr r t 1 tr 1 ré é ts é ér t sés s r s é té t tr ét rt s ss s r s q r é t r t r t
31 s s r rés t r s 1 ét s tr s r t tr r s s rs tt t ès t s t r s rés t ts ç s s tér ss r 1 rs t s rt s r tr s t s ét t t s q r r èr ét s r r ér t tr s è s t t t é à s é s t t t s 3 r rs s ér é s s q s t tés 1 r ss 3 s 2 r s â s sté r s P r q st 1 è ét s s t s s r r t t 1 r ss s t s t r s t t st t 1 r ss s r t r r r rt à 1 r ss s P r s ét s s s é é étr t r t éq st t st s éq t t st q s t é s r P r s q s r s r r s r s st t st q s
32 tér t ét s r s s s s s t rs 2 r s r s à é s r s s s s t sé s é s é s à rt r é é ré é s t r é s r s s s q tr é s s s 3 té s q tr é s s s r 2 té s q tr é s s s é t Pr s 1 î s r s 2 s té s q tr é s s s é ô r té s r s s q tr é s s s é t Pr s 1 î s r s 2 s té s t t s é t s s r s s t été é é s t à s té trô é s r 1é q t à 2 èr t r P r q é s s 1 t s ré ts t été ré sés t t t â s t t s 1 r s P r t é s s q tr r s ts t été ré sés q s tr é ér t t â é ér t s
33 tr s ré ts r r s t s s s t q tr t s 2 r s r q t s s 1 ér r t t s q tr é s rr s t à s ré ts q s r t tr t s s s t s tr t s 2 r s s s t sé s s r é t s ré ts P 1 ér s s r s tr q tr t s s s s t q tr t s 2 r s â r s s r s s s é s r 2 2 s 2 r s t été t s r s t s â s s s s s s q é s s ét t q é s r s t é s s q t s â s ss s q é t été ré és à é r s s tr s t s ré ts ér ts st t r s r s s s à r t rs s t t t 1tr t s t rs s r s â s t été és à s s s t t sé t s r 2 r 1tr r r s t r s r s â s t t s r µ r t r 2 r t s r s s s t été rétr tr s r t rés é t s rq és P
34 P s à t 2 r t s s t sé s s tt ét t r ss q à s rs é r s s t s tr s r t s t r q t s r s s r t s s é ss t r s t str t t s t r t té t q st t t s t sé r s t s é ôts s t s é t r s t s é t s t s t r t t r s t t s t s s r 2 r t q s t 1t t r s é q t s s s ré s r st r r 1 r s s t s st ré 1 q rt s ts t st r s ts s té s é t s s ts rt t étr r à r s rr r 2 s2 t t s r s s t té s s t s s t été és s t r és t é sés s r r 2 q r t é sé t t à s t r t t r q s r r à r s r r q t té é ôt à q s t r èr 2 r t s s s t 2 r é s 1 s r èr 2 r t st ré sé s rq é P é t r séq t r q s q é s tté t t r t t r s st r rt à q t té à q s t s 2 r t t r rès és 2 r t t s 2 r t st ré sé s térêt t rs r s 2 r t 1 r s t r é r r s t été é r str t r s s t r rs s s s ts r sq s s 2 r t t r ét t ér r à q s é s té s é t s t été é és 2s P r s 1 s 1 r t s ré r é s s ts s été s str t s tt ét st s t tr rsé s t s r t s rés t ts t s s ré éré s r r 1 str t s r t r s t tr s s ts à tér r
35 Valeur d'expression A M r ér t s r s t r t ré rt t s é s r s s té 1 r ss r tr r r t 1 t st ér s st s 2 rès tr s r t r t q s s r ré rt t s é s t r r 3 t té éq r tr s s 1 q été ré sé s t r q s t s 2 r t 1 r s 2 r t t r r èr ét été r s t tr s 1 s t s s r t à s q s t ê rés t t s r s à q s t r é s s 1 s r t té r é s trô s s t s q té r s t été s t ér é r t t r ît r s s è s r sq s q t q tr é s s s r s s t été é és 2s 2s st t st q 2s st t st q st sé s r s ér t s s s t t r st r r q è P r ré é r s è s ér t t 1 r és s s t sé 2s r r q è rr t è t sé st t q Y ij = µ i +E ij, ù st t s s s t t s 2 r s st ré t q Y ij st s rès tr s r t
36 Effectifs P values r str t s s t st é é té r r t q t r s t µ i st 1 r ss 2 è r t t E ij r té rés è s s r à t t s s t s st é 36 9 = 27 rés rté r rt s è s q q s è s t s é s q t s s à r s t é é té r tr r été ér é t st s é st té été r é r t s s è s 1 tés 1 1 s t s s s r2 t r r r str t s s s s ré sé t st é té 2 tr s t s s s t t s t tr s q tr t s s s t s s r s s t été ré sé s s t st t s t 1t è q r s r s s 2 r s s r ts s s ré sé t st r s tr s 2 s q t 2 r t s s 1 t s r t s é t t é t 2 s t s 2 r s t 2 s t s r t s P r q r s s s r t s t été sté s r ét trô t 1 1 s t s r t r t s t
37 étér s s P r 1 r éré té s s 1 é str t s ts 2 r s t s t r rt d/a ù d st ér 1 r ss tr s 2 r s t 2 s 1 r ts t a st t é ér 1 r ss tr s 1 t s r t s s t q t s s r s t t r t 2 d/a = 0 rs 2 s t 2 r d = 0 0 < d/a < 1 2 t 2 r tr s r ss d/a > 1 2 t 2 r tr s r ss rs s rs r t s s s ré sé tt 2s s t s è s ér t t 1 r és tr s r ts é t r t t s sé r é té 1 r ss tr s r ts r s s r t ét t r r s r s s s t s r r s r s tr t t t s s t sté 1 st 1 ès è s 2 t r s s r s t r r rt à tr P r è g t σ 2 g,a t σ2 g,b s r s 1 r ss é q r s t t s s t s t R g s r s ér r à r r t é s è s s s ù s r s t s s r t r s rs q s t tr s r r 1 r ss q t rs r rt R g = σ2 g,a σ 2 g,b s r s ér r à r r rt s è s t s r t st s r r è s N AB 1 r sq s r t r q R g r è s r s ér r à t p AB r r rt r rt t é r q s t st t st q q st r s è s s s è s r sq s σ 2 g,a > σ2 g,b t st H 0 = {p AB = 1/2} r s t r s tr H 1 = {p AB > 1/2} r 1 r ss s ér r 3 N AB 1 s t str t B(G,p AB ) r è s t st st rs P(N AB 1 > n AB 1,obs p AB = 1/2). tt st r té s r r n AB 1,obs è s r sq s r st s ér r s t s s 2 t ès H 0 s s q 2 s è s q t r s r s t n AB 1,obs ét t r è s r sq s t t s tr é t σ 2 g,a > σ 2 g,b
38 t r é s s r ss t rr r s 2 t ès tt 2s r t r é s s s r st r q r s s t é s s t s ér ts r s é s r q t t r é tr t s r s r r t r é s s r s r s s r s s q tr s s t sts ét t s t s r é s t s str t s s t r séq t rr r r èr s è r r s 2 t ès t r t s s ré été s s t s s s è s r rt é rt t2 R g = tr s 1 t s r rt s ér r s à s r tr r r s r r ss t st t r é s ét t r s t t r tr s t r s ré été t st t à s r tr r ét t ér r s s é s tr t s s s r é s t sé s tr q ss t st t très r t r é s q é s r r t é à très ss t s è s s s ré s è s rés ts s r é t s t t ss t rr 3 t q 2s s t r s t 2 tt 2s ré é é r 1 s s s t r s t r r rt é q r ss t t s t tr ré ér r s 2s s s st s è s ér t t 1 r és s s s t ré s st s è s ér t t 1 r és r è s ré ér s st t é r s s è s rés ts s r P r 1 r r s t s t r s r ér r t s è s rr s ts s s t sé s é t 2 t 2 s rt s r r t 2s s t é s r tt ét s é s ét t r s t r s é r t ç st r sé t é r r ss s q t s t r è s r r s s r r rés t t t r s t s st è s ér t t 1 r és s t r s ét t r sés ç ér r sé é ér t s rs t r s s r r rés tés rr s t à ê r è s s s t sé s r 1 t s s s r 1 r s r rés t t s t r s ss
39 Effectifs Effectifs P values n = P values n = 5 Effectifs Effectifs P values n = P values n = 50 r str t s s t st s r s r s t s é s s é s t é s r st r r t q tr s t s 2 s ér r s s é s r tt str t s s r str t st s té r r é s t sé s t rr r st s s
40 t rr 3 t P rq t ù s s é tt 2s t s r tt t r r st è s ré ér s r s r r rés t t t ss t r t s st s t s t s rés t ts s t t t r rét s t ér ts tr s s t stés t t t st ré èr t s à r s q s s é tt t s s s é s s s 2s é t t s q 2s ét t t é à t ré s r s é ss r t s rés t ts r t t t q s 2s ré sé tr rs s s s é s s st rq s sséq s s rés t ts s è s 1 ê s t r s t s 2s tr s é q 3 s s r séq ç é ér t t s é t s s s s s t été té s s r t t t r s s té r r é t r s s r étr t s r r rs st t t t r r q s r s t t r rès s é ô t t t t tt r st t é q st à r tr té t st r é r s r s ss q s q s ss t s tr t ts ré rs st très r q t ré é à t r t s t ré t t s s à 1 s t 1 st s s s té tr s é t s î 2 tt s t é tr s r t ôt r s r èr à t r tr s tés é t t t t t tt t été ré é s r t r t t ré à 2 tt s s té rr s t à rêt s r rs é r s é r é s r rs té é ét t s ré è ts t été ré sés à è s t r t t s r r ss r t r r t r r 2 tt s s é r q t é s s s s r r ss r t r s r st r 1é q é r s s ts â s r èr é ér t t r r s 1 s r ê q t r st à r r é t rès q rs s â s t été és à s t s â s
41 ss s é s ér t t r q t t r é é s r 1é q t r é é s r P 2 tt é é s r 1é q 2 tt é é s r 1tr t P r q t s s ré sé q tr 1tr t s ré q s à r t r r â s s s t sé t r 2 s r 1tr t tr t t q té t été ér é s t s t à s r r s t t é tr rès r 1 r r s q tr 1tr t s ê t t rs été r r é s r séq ç P r tr s rt s é t s t été ré tés s ét Pré r t r r t séq ç ré r t r r t séq ç t été ré sés r tr r s t rt r s é t s t t s 2 t été sé t és r s2 t ès é t r été s2 t ét sé t s t r é à t tr s r t s rs r s2 t ès r r r s t s ré t t été s s t èr q s r s ét t t s t é tr t é t s P r séq ç 1 è sé t t été t é r s ré r t r s t r 1 r ts s r t tr t é t s q té r r s t été ér é r é tr rès r 3 t séq ç s r ts s t été ré sé s r séq ç é ér t 23 r 1 s s str t s r t s s s t s r str té t 1 r ss tt t q sé t s 1tré tés s tr s r ts r séq ç q t r r q t t r r rt à séq ç r ts é t r s r t s tr q r t r s r s séq s t r t 2s tt r èr t t q s r ts séq és ét t tés à rt r s séq s s é t s t r ts r s t t t r s r t s r s t s P t s séq s t r été tt 2é s
42 t t st q s séq ç t rt r r s q tr é t s t P t r s r t s t r s r r s ts s r ts q s ts t s P s t ts s r s 1 ts q s ts t s P s t 1é q t r P P t r 2 tt 1é q 2 tt 1 rt r été ré sé s r s s q t r s t r s 2 t s êtr rt r é s s r st r té r P s t st r r q r t r t s t t rs séq ç t é t s r ts tr s q té rt r été ré sé r t s s s rt r r s séq s r s r é st r s r s tr s r ts r sq s t s é rés t t s r é s s 1 st s té s rt rés t t s é q tt ét rt r été ré sé t s t r r t s t s séq s s s r t t r s t sq à 1 t t s t 1 rr r t tr s st t st q s séq ç tt 2 t rt r s r s 1 é s s é t s t s t été r sés r r t r s t t s r s rt é é ts tr s s s r P P r è térêt 2 è t tr ét é à s ttér t r r t s s 1 é s réq s s rt é é ts tr s s s r P 2 r s t P P r s s t sé P tr 1 2 r s t é t rés é é t tr s s r r s s s t ér é t s s étér 32 t s s q s 32 t s r séq ç r
43 SANS INSERTION Site d'insertion potentiel de Juan 300pb AVEC INSERTION 600pb Amorce sens Élément transposable Juan Amorce antisens interne Amorce anti-sens r ét t s rt r P tr 1 tr s r s s s rt s t s t r s s s rs q s s rt s t s t é é t t r
44 2s st t st q 2s st t st q été é ré r r st r2 r t q s éq r P r s t t st q s t é s s 2s s st t st q s q é s 1 é s séq ç é ér t s t é ér t sé s s r t s t P ss s r s rsé r t s t s 3 t s r é r s r t t à s r tèr s r t t s t r tèr s r s rsé s é s ss s r s rsé r 1 r ètr s λ 2 s t s t Φ r ètr s r s rs t t r tt t t s r ètr s r(x) = λ Φ s é ss t t é ér t s ré ts q s s t r r t t r s r s rs r q è s s s r s r s s ût s ré q r séq ç st 1 q s s r r tté st r r s s û t r s 2s s st t st q s t tr 1 ér s s t é 2s 1 ét s s s s 2 t ès s s t s r r té s è s st s ér t t 1 r é s è s s 1 s r s 1 r ss tr s q tr t s tr t ss ê s r s rs s r r t s 2s è r è été ré sé t s t è s t P ss s r s rsé X gi P(λ g,φ g ), X gi 1 r ss è g r t i λ g t φ g r s t t 2 t r ètr s r s rs ss és è g s è λ g é s t q st rt t s s 2 t ès s ér t s t s t rs êtr t sé s s ré ts q s r t r st t φ g r ètr s rs φ g P r s è s ér t t 1 r és r st st é s s s rs q st s r st é r s è s ér t t 1 r és s 2 t ès st t s r st φ g φ g t êtr t t s t st t r ss ét st t 2 φ g s r s è s 1 r ss 2 s r r tr r ètr s rs t 2 1 r ss s è s s q r ss st t φ g t ss st 1trê t r t 2 ré 1 q rr s à r t tr ô tr 2
45 r st t r ètr s r s rs φ g t 2 1 r ss λ g é r t q q t rr s à è s r s tt t r rés t t r s t t ré r ss ss t st t ré r ss 2 ré 1 r t st r ré rt t s s r r s s à t tr 1 ré rt t s s s s r s s 1 à 1 s t s r rq é t rs r t à s r st t r ètr s r s rs 2 t r séq rt ss s s t r rr r t r P ss s r s rsé q st sé 2 t ès rs r é r s t r t s rs t r s t s s 1 è t s s s t sé 2s è r è t s t s P ss s r s rsé r t t r ètr s r rs st é à r èr ét X gi P(λ gi, φ g ), ù λ gi st 1 r ss 2 è g à t i t φ g st r ètr s r s rs st é à ét ré é t s s s t t sté s s s é r s r ètr λ gi t s t s t sts r rt r s t s s t r r t tr st r ré rt t s t st H 0 : {λ gam = λ gbm } s H1 : {λ gam λ gbm } t s r r é s s rs tr q r èr ét tr 2s s r st r ètr s r s rs r s rés è s ér t t 1 r és tt s r st t ss tr t st r ét t r s è s ér t t 1 r és s t s rr r 2 rs t r s s s t s s s t é rr t ss q r s t sts t s
46 q trô t 1 1 s t s s s r2 t t r t s è s s s s s s st s è s ér t t 1 r és à r r s r r rés t t t r t rs s t r s s à ss t sé t ss t rr 3 t s s é t 1 é t s st s è s à r r tr s ét tés r s t r s t s t 1 t ss r à ôté t s rré é s t t 2s r st rt t s t
47 és t ts t s ss r s s tr t r s é q s t 2 r s s s à 1 ér t sé r s r r t r s s s è s r s s t s t r s s tr 2 r s t r ts P r q t s s t sé q tr é s r rés t t q tr ré ts q s t s s s s r és t s r s é s é s ré t t r s ts é t r t r s rés t ts s r t r rés t t s s t s ét é s t s t s é s rès t t s s é t s è s t r s t t 2s st t st q s s é r 1 r ss r è s s t r q rt é r s s ér t à t s t s étér s é q s t t t tt ét rés t s t s t s rt t s séq s é t r s st r r r r s s è s r s s t s s t r séq tr s s s t térêt t ré r s s s s s r s 1 r ss t s t t t é é rt r è str s t s s t s è s r q tr s è s r t s s t tré sé èr s s 2 r t s à r tr s t térêt t q r è s t rt t s s t r r s è s r q été ss é tr s t t ss tré q s r 1 r ss s è s r s s s è r s t s rt r t s r s r s s s t s t s t s s st r tt r èr ét t s q é s r s s st r t à t q r tr st r t s s ôté
48 t st r t s tr s t éq t r tr s s t s st té r s rs q sé r t s 1 s è s st r st st é à 1 à tr s s é s s t 2 t r tt t sé r t s 1 s è s st t à 3 s s ré t q r sé r t st r q s r t s s r q s st té ç s r r s 1 s è s s t r s s s s s s rt t s ts tôt q s é t s q t s s t s s t ré st r q s 2 s st t êtr é té à t t s té st ss r q rt s è s s t rt t tés r 2 r t t r s é q s s t é s rs ér t s râ à s s r ts t s t r r t P r s t é r s 2s s st t st q s P r ét r r s r st r st r ss t s r 1 r ss s s r r é rré t tr ér r séq s tr s 1 s è s t st r t ér 1 r ss 2 tr s 1 s è s rt s tt 2s st s r t sq ér s 2 r s rt é r s s t s t r s é q q s è t st r èr t s s ré sé tt 2s s r è s 1 s s r sq s rt ét t té q t é t ré r s s s t 1 r t s t s séq s ss s s s s é s tt 2 s r t s s s s r é rré t t rré t r ρ = 0,0054 s s t s r t s q st r r tr s t s s r t s st r s s s q r t st r s t t q s 3 s t s s r t s s tt ét s t rs t s s r é s tr s t s s s r s r 1 r ss t s t s s ss é s à rt r st r 3 2 t t tré q r séq s r t t r r q t à t ss t st s s t r rr r s s t é r èr ér t s s s é ér r séq t st r s s s s st r s s s s t é r è s ér t t 1 r és tr s t s s r s è s s à t t s s r s s
49 r r r rés t s q t s str t r r rt à 1 tr s ss ér r st r s s s s st r s s r è s r 1 t r é ér r r è s r s è s ér t t 1 r és tr s t s s à r ér r s r é t rs s è s ér t t 1 r és s s s q q s t t t r s t è s ré é t é r t q s t ér r 2 st s s t t ér t tr s è s ér t t 1 r és t s è s r s é t r t t 2 t s té à ss r tt 2s s s s t sé r q t s t r r r s 1 str t s r tr s q t s str t r r rt 1 q t s tr r s r très ér sq à r rt r 2 é t r t s è s ér t t 1 r és t é t st s s rt rsq r t ér r st s rt r s è s ér t t 1 r és q r s è s é t r s tt t s t s r tr s s t s s ér r tr st r t s s ôté st r t s tr s r tr t è s r s rés t t 1 1 t s ss s r t êtr û à r séq s q t r t 2 r t é ér t q q s 1 s t s s t é t s 1 q r r s r r
50 1 r ss s è s q t ss s r é séq s s èr q t s s è s ét és s t ét s t s t és s rt sé r q st à r ér r ér r à rés t t st é s s r s s q s ssé s à t s r s t q s séq ç é ér t s s rt tt t ès r s s 2 r s tr t s r t s t rt r t 1 r ss 3 s 2 r s s s ré 1 r ss 3 s t s 2 r s è été s éré ér t t 1 r é s t s t s ét t ér t s r ts t 2 s r ts è ré é é ér t t 1 r és t 1 r ss t r é r tr s 1 r ts s s tt s à tr r s ts ss 3 rts s 2 r s ét t stér s t t 2 t s rt2 t t ré é é s s è s ér t t 1 r és tr 2 r s t r ts 3 s ê s s è s P rt t s s s r é q t è s ér t t s r 1 r és 3 s 2 r s r r rt 1 r ts s t t s s 1 r é s t s t t s s r 1 r ss q s s 1 r ss t st χ 2 à ré rté = 3, t s è s t été ré é és s 1 r s s q t q t t r r ç s t q t t r 3 é s t s 1 t s s s é é s 3 r s s t 3 ré rt t s s è s rt t s s 1 r s s P rq ss ér t s tr ét r r rt à rt2 t q t ré é é s s è s P s rs 2 t ès s t 1 q r tt ér rt2 t t s ér é s ts t s P t êtr rt s rs è s t 1 r ss r 2 rs r ts ss té q s s 1 é s s t sé r rt2 t st é té r 3 t rt t é é 3 r s è q rr t r s t t s q t ér s rt t é à sé r t s s t s 3 str r à st r é t t r t r t s s ss s r 1
51 ét rt2 t 1 é q t 1 r ss t st r r r t rés t 1 t s s t s 2 r s ét t stér s t t 2 t st r q 1 r ss s t r s t rt t rt r é t t q s t st s s t tr és t s s r t 3 ï s r és 2 t rè é ér s t st s tr t tr 1 r ss 1trê t s é q t t tr r s tr t r ét rt2 t r tré s ér s q s r t s ét t s s tr ét s r r s t r tr t ét t 1 r r q t r t s s r r st rq s s s s tér ssés r s t r ê ét t r tré è s ér t t 1 r és s r és s t tr 2 r s t r ts ét rt t s r r t t s s tt ét s t sé rr t r s t sts t s s t st t s t s 2 è s ér t t 1 r és tr 2 r s t r ts s r s 1 és s t r rt è s ér t t 1 r és s t t s ér r à s r é r t r t st χ 2 à ré rté = 2, è ér t t 1 r é st tr tt ét t ôtr s t st t été ré sés à r t 2 r r s r s t sts q s r t s t s 2 s t è s r s r rs 1 r s s 2 r r t s s r t t t è s ér t t 1 r és s r s 1 r s s s t é t è s ér t t 1 r és r r rt à q st tt r s r t 1 q r rt ss à t q q s t s s 2s st t st q t sé rs r 2s st t st q s rr t r q rs é s é t r q è s rt rt t s t à s s st t r q st r q s t s r st é r q s t rs rt rt ss r t t é t ré s 2 r s s r ts s r r s t r 1 t t t t té s tr 2s t à ss s t s t sé rr t r s t sts t s ss t s r r 1 s t s r 2s ré é é è s ér t t 1 r és s r q rt t q st s s t t ér t s è s s r és s tr ét s rr t r t st t t st χ 2 à ré rté = 0, 16 2s t s s rés t ts t s q s ét s t s t r s t r
52 è s s t t s r 1 r és 3 s 2 r s r r rt 1 r ts t s t r s q P t s r t s r s r s q s s 1 s s s s 1 s s r P r s è s ér t t 1 r és t s r r rés t t è s és r s r r s r t s r s s t 2 è ér t t 1 r é tr 2 r s ss s s t s 2 t s 2 s t rs r ts ré è t ér s 1 r ss t r r t r s r t rs q ét rt t s r s t st s rt2 t ré è rt s rt r t s 1 r ss s q s rt r t s 3 2 r s t tré s s r s t st s t t t s r s è s és à s r t ès t r r t tr t r q 1 q r t s ér s tr 1 r ss s r é tr s ét s è s ér t t 1 r és tr 2 r s t r ts t tr s è s ér t t 1 r és tr s t s 2 r s t rs t s r t s 2 t r s t t q tr t q è s ér t t 1 r és tr 2 r s t r ts r s t s r t t r s t s 2 t s 2 s s è st à s 1 r s s è q é s ré t tr rès r t st q é s t tr rt r t 1 r ss q t êtr rt r t r t s t é s s q tr st q é t st tr st é t q é s str t t t s t t 3 s s r t 3 ï s s rt r t s s t tr tr î t stér té t st t rô s t tr s tr r t s tt ét
53 s t rs t tré q s r 1 r ss è tr r t str t r s r s 2t s t r st t t s très s é t t t q rt r t 1 r ss s r é s tr ét st rés t s s t 1 t s s r q tr tr s è s ér t t 1 r és P r s tr s è s ér t t 1 r és s t èt t 1 tr 1 s t r t s s é é 2t sq tt t t rô t êtr é rô ê s r st t t s é t 1 è s s t r rô s é t s2stè r 1 s t s t è r st q é s é s â t st 1 t t r rô s stér té 2 r s t rr t è st t s t2 2 tr2 s s è s ét t ér t t 1 r é tr 2 r s t s è s r t s s ét rt2 t s t s è s rt r és s r r rés t t r s r s t è s ér t t 1 r és tr 2 r s t r ts s 1 s t s t és s r r s s t s t t s q tt s s 2 t ès ré rt t è s r s r s r s q s r r rt 1 è s t st s r 1 t 4, s ts 1 r ss s r és s t s ts ts 3 2 r t t s r q s s t é étèr s 3 â stér té s s r é t 3 ét t é ér t rt s tt 2 t ès tt r é t à q s s s r é 1 ès s t t s s r r s r t P rs s s r t s s rt t 2 t ès ts t st s s è s tr s 1 s 1 s tr 1 r ss st éq t t é r t st r r r été r ss s s é t t rs rt t é tt t rt t r t t st rs t t s 2 t Pr s r s Pr s r s s t r s rt tt t é r s è s és r s é t 2 s r t q s è s t s 1 r t à s sé t s s r é 32 t 3 s â s t s r r s ét s q s è s és à s t sés rs s 1trê s s s t é ér t s s sé t q s t s t é t P r 1 s è s à 1 r ss t t t st r s t s s rt sé t s t s t r s rt s è s és r s t é t rt èr à r rés 1 t rs t s 3 s s t t rs 3 s â s s q à r t
54 t s t r r tr r ss r t t s tré s té s r t rs stér té s r q s r s t s s rés é 2 s r r rés t t r s è s t 1 r ss st rt r é 3 s 2 r s s r 1 r ss é t s r t r séq s s è s és à t t t 1 és à r r t â t à 1 r ss t st r s té t r stér té è tr s ér ts r s s t s s t r 2 t ès rt r t s t t r s rt r t s é s à s t r t s tr r s t s t s s 2 t ès s t r r 1 tr 1 s r stér té 2 t r t s 2 t Pr s r s ô ré t s tr ét tr s ét s ré é é très rt r t s 3 s 2 r s t r s é q s r s t r r t r s r t q s éré t tr s st à r sé r tr è é P r rs s s tré rt s r s 1 r ss tr s è s r t s r s t st ér t è rt s tr t s s r ré t tr s s t é tr t tr t s s rt s s r ré t s q r ss st é ér t té à s s tt s r t st r é r tr s ét s tt t étér s s s s 1 é s tr s étér s s s s s s tt ét q s è s r sq s 1 r ss ét t ér t tr s t s r t s P r s s t sé r rt d/a r ét r s è s tr t tr q s t t étér s s à à t 2 r é r 0 < d/a < 0,5 è s tér r t és és tr t s t 2 r r tr t t 2 r tr s r ss rt 0,5 < d/a < 1 t tr t t 2 r tr s r ss d/a > 1 r s tr s étér s s s é s tt ét s t t r é r s tr t 2 r très q q s r t q êtr s r é s rt s ét s s t rt2 t t étér s s rt s r é s 1 tr s ét s s t
55 Nombre de transcrits France Zimbabwe Nombre de transcrits Seychelles Kenya d/a d/a r str t s r rt d/a r s q tr r s ts 2 r s ss s s q tr t s s s d = 0 2 s t 2 r à s ss s r r ss t s étér s s à t 2 r tr s r ss ss s ss s s t t 2 r tr s r ss s rr s r s rr s t 1 s s t
56 tts t t rr s r s s t 1 q r s ér s s t r s s t t été très ét é s r tts t t rr t 1 r s é s r t s s s é s s s r t sé ré s ré t s s étér s s s r t rè s t t 1 s té s é s r t s t s s s t rt t s s 2 r s ts 2 r s tr s r ss s ét t sé r ét r r étér s s q s 2 s ss 2é s r r 1 1 ét s s s s s ét s ré é t s st ss q s ér s ét q s t s r s rés t ts t s t r s t rs r s rt rs ré r t ss s s s s ré é t t é t 2 r s s s 1 s s ê s s à r s à r t té s s rt t é t s s ts r s s è s t s ér s s s ré 1 r ss tr s t s s r s q tr t s s s s s ré é é t è s ér t t 1 r és r s t s r s t t r s 2 s t 2 t P r s è s s t s à t t s s r s s t s r t r s t és és è s ér t t 1 r és tr s t s s s tr s t s r s tr t t t è s ér t t 1 r és rs q t r ç s rés t s t t s t st χ 2 à ré rté = 1, tt s r t st r str t r t s s s r é s r s séq s t s t r2 t r s t t s ö t ött r r t s ét s t tré ér t s t s r é s r r rt 1 t s 3 str 2 à s r s s t é t tré ér t s t s r q s t t s r tr s s tt ér t s r s s 1 r ss s s s rs q r s r t t s r P rq t s éré ér é r s ét s séq s s rt t 1 r ss s t s 3 str st q st r t r tr
57 r è s ér t t 1 r és tr s t s s r s ér t s t s s s P t t r 1 r és a s 1 r és b 2 2 s r s a è s s r 1 r és 3 s s r r rt à s b è s s s 1 r és 3 s s r r rt à s tt t t s 2 t s 2 s s q t ss r s r q t st s t 1 r ss r t t éq t 1 tr s t s r s st s rt t rs s r s s 2 r s r s t q tr s ér t s 3 s 2 r s r t r ç s t tr st t s t s r q st s s ré tr st è s ér t t 1 r és s è s ré é és s tr ét r t s s t s r t s s è s ér t t 1 r és s r ét ét t rés ts s r tr s t è s P r 1 è s s t é t rés ts s tr st è s ér t t 1 r és s ér s t r s rs 1 t s 1 s é s s s t sé s é s ré t té s rs q s t t sé s é s tr t s t s r t r ss s s st ss q 2 t ér ss tr rs s t s ôtr s s t ré é é è s ér t t 1 r és s r s r s s r s t ér s t t st χ 2 à ré rté 2, t s s s s t s ss t s s r s û r tr r rs è s ér t t 1 r és s s 1 s ét és s t tr é s r s é s s t â s rs q s s tr é s r s â s q t t rs s ét s t tré q s 1 très rt s r s tr s 1 r ss t rt2 t t r t
58 r s t 2 s r r rés tés s s è s s r 1 r és 3 s s r r rt à s q q s t t s s 2 r r t t tr rr r t t2 P t s P r t s s r r t r s t é r P r ss s q s t r s t r s s t s r r é r ss t s t té r s r r rés t t s s r r r s t été é s é t r q t 2s t s s ét é s è s ér t t 1 r és t s t r sé s r s t r s t 2 s s 2sé sé ré t s è s s r 1 r és 3 s s t 1 s s 1 r és t rés t s t r s t 2 s r r rés tés rès ss t r s è s s t t s 1 r és 3 s s r r rt à s q q s t t s s r é s s 1 é s è s rr s t à s t r s s tr st è s ér t t 1 r és t é r tr rr r t t2 t s s t s r s s r r t t r s s ré èr t à s è s s r s 2t r s P tt s r st q é t t s s r ss s ét 1 t s 1 tr s t r s t s ré èr t t tr 2s à q è s t tr s s t rés ts s tr st è s ér t t 1 r és r 1 2 r s r 1 2 r s t ét2 tr s ér s t r r è r s t t s s è s és és r t r s s s ét té ér s 1 r ss s r s r t Pr t s tr s è s t és s s ét s ré é t s r t r t ts t t r t s s t s rés ts s r tr t r 1 r ss s êtr 1 é s tr s è s s t r rô s é rt èr s
59 2t r P r â t sé t r ét 1 t 3 s ér t s ét s s s t tér ssé s à é ét q tt t ér t tt ttr t à t t s tt s ré s s ét s é ét q t tré q rés st ét t t t q ét t à ér ts t rs ré rt s s r t s s r s s 1 tés s 1 r s s très r s è s t s ts t t tré q s t ét t q és s ré s s tr s s è s r s s tt s à tr r s è s q és s s é è s r s è s ér t t 1 r és tr s t s s s é t s è s s ét t ér és rès s é s t s t s r é t r rt r t é rt èr s P r s ét tr s r t st s ss st r r s è s ér t t 1 r és 1 q s r t s ér t t 1 q s r t séq tr t t s s ét r r q s è s t été r t t q és s sé r t s s è s t q s è s t r é rès r s r ss s s s st st q s P r s è s s t t s 1 r és 3 s r r rt à s s tr s t 2t r s P s 2t r s P s t r è s sé r s t t à t è s t s 3 st r 2 r t t s r s è s 3 s s t t s r s è s 3 s r t t 2 r t 3 s é s t s 3 s s sé r t s s s s t 1 r ss è s ét 1 t é r s é s t s tt q r t s s r rô s rt t tt r èr s rt s è s q s t 3 s t 1 r ss è s s 2t r s P ss été s r é r r t s s t r rèt t tt t 1 r ss séq s é s t s s ss t ç très str t à t r tr s r t ré t r été t 1 s 1q s st 1 sé r â t s r ss sé t s r rt s è s ét 1 t s 2t r s P s ss t t tr s ér s t r s2 tr s ér s 1 è s ét 1 t s r r s s r s s s s ét st ss q s s r t s s t é s à s é s t st ss ss q s s t s à tr r tér st q s q s t
60 s r t s é t s ér s ét t st t s q q s t t s s q s st ré st s r q s s t st t t s t sé s r é t s rs s é s t é t r P r s è s s r 1 r és 3 s s r r rt à s s t r s t s és t 1 à tr s s rs s é s t s è s q és s s ré t s r s t t 2s t r é r t t s q t té t êtr st é s t 1 é r t s rt 1 r ss 1 3 s s q s q t té P r s è s s r 1 r és 3 s s r r rt à s s tr é t è t r r r t q s rô ré sé t é r t tr é t è t r 2s r 1 t r t s r s s 2s è rô s ré t s st r t é s é r st s r rô ré ér t 3 r t t r ré t s s t 3 t t r t ès t ès r t t s t t s rô 3 s â s st très st t r rét r s ér s s r é s r 2s q s é rô s t s r té s sé s s s ss r s r t t rô êtr r é r ét sé s r s t ts s t s t ts t ré t té à st é tr t r t r té s q t rt té â s t ts tr t é t tér t rt t r q s èr rô s rt t q s t r t t r t t rt r t s2 t ès r té q s s s ss r s t é 2 r s t rs s é st s ét 1 t s 2 r s t t 2 r s t t t s é té s s t t t s 12 s 2 r s s s s té s à r é st ss q s s t q é s s tr r ét s t r t t st ss q s è s s t é t q és s s ét 1 t s 1 è s q és s ét s s t s r 1 r és 3 s s
61 r ét s t ré s r s 1 32 s t s è s s t s 1 r és 3 s q 3 s s é t rô s r èr ét s2 t ès sér t r é rès r s t r r rt à s è ét2 tr èr s t st q é s tr s r t ét2 r r s t è P é r 12 s tr s r P t tt 32 t 2s é t r èr ét ré t t à sér t s r t 1 è s és ét s s èr rô s ér t tr s s è s q s t t s é t t ér 1 r ss s 1 è s t êtr é t t é ôté à r t sér t t tr à ét2 3 r s t ré t rt té t t té ré s à rt r r s t t ï t t r t ss t é r r s t é t rô rt t s r r t rô é r t 3 ré t s2 t ès s ér s t st t r â t 3 s â s rô s r r â s â s t r s rs t 2 r t rés é s st q é s r 1 é s s s s t r très é r ts 3 â t st t r rét r s ér s s r é s 1 r ss è s q és s ét s
62 r è s ér t t 1 r és tr s t s s s s s s ré é é è s ér t t 1 r és tr s t s r s t s s ré é é r s t t t è s ér t t 1 r és tr t r t s t s 2 t s 2 s t r séq s rt t s t s r r t s ét 1 t t 1 s s r t ss s s ér r s t s 3 s s ér t 1 r ss tr t s s s s s 1 é s ér s 1 r ss tr s q tr t s s s 1 t s r t 3 str s è s t t s 2 s t t 2 s 1 tr s t r s t s t s ér é s rès ér t s ét s é ét q s t s t s t s r s t t s ö t ött r r s séq s t s t t r2 t r ré t r è s ér t t 1 r és tr t s t t r s s s s r é ér s 1 r ss tr s tr s t s r s
63 è s ér t t 1 r és tr t s s s r > r < s s s tr s r s trr ts 2 s è s ér t t 1 r és q tr t r ç s t s t s r s è st r tr é s s tr s r s s s s rs s t rés ts s 1 r s s s r s s t r ç s t ré é r s ér s 1 r ss r s t t t è s r s t s 2 t s 2 s st très r s t t t P rq Pr è ss 2s st r r t s t 2s t q s s ré é é tr t è s ér t t 1 r és tr s t s s t r r à s r t q s s r t q r q rt é s r r t s ér s s t rt r s r s s q tr s s rr t r s t sts t s s r t s str t s s tr r s ss s ér s tr s t s s r t s t s t r s s à rt r t ss r q s t t ré é é è s ér t t 1 r és rr t r s t sts t s s r t t è s 1 és s t s t t s q s tr ét t st χ 2 à ré rté < s s t r 2 t t ré 1 r ss 1 é s r s r t s r s ér t s r é s s t tré str t r t s t s 1 r ss è s ér t t 1 r és t rs rés t ts s t t rs tr rsés s t t té s ér s s r ss s rt s t s é s r s t sé s s tt ét t é t s r à q t 1 r ss é s r s st r t 1 r ss s r s q st ê rt t r s s t ss s s t s r t s s s ér t s r é tr s t s s s st s r s s r q s s s té ss r s 1 r t s s ét r ss r rt r é t s r r r q t t
64 P r s è s s t t s 1 r és r q s s t r s tr t q tr 2t r s P t tr s s r tr t s 1 r s s s è s t rô s ét 1 t st ss q r t t r ç s t s s à s rt r ss r té s r ss s ét 1 t r r t r t s q t s s st s t s s t s q r q t s s s s s é s r s ts s s 3 s t s t s s q r r tt 2 t ès è st é t s r 1 r é s t r ç s r r rt 2 t 1 2 s s 1 t s r s r s 3 str s r t ér t t ré é tr s t s t r st ss q tt é r t s t s t s rt t r t rq s t r s t r r t r tr s t s s è s és à r r t é t r t t t t s s 1 s 1 s s s t sé t s 1 t t st s s r r t s r tr r s s r rs è s ér és tr t s tr s è s st s é t r è s s é t s r 1 r é r r r rt à 1 t s 2 s t è st q é s s r t ès t st é ss r à ét ss t é r t r r è rés t s 1 r s s st s s 1 r é s t r ç s r r rt 1 t s 2 t s 2 s P r s 1 t s st s è s s 1 r é s t t t é r s r s s s q s s sq s s r t q é s t s st r s s t r rét t s ér t 1 r ss r tr t ér s é s à st r s t s s s 1 é râ à t st r 1 r ss s q tr t s s s s q t s t r t r ç s st s t t s é é q t t s s tr s t s à 1 t t < 0,005 s rr t rr t s 1 t s t r s é é q s tr s tr s t s s tr s r s s 1 à 1 t ré é é s ér s s t s s s très r s tt t tt s r t st s tr t r é é té r è r è t sté râ

65 P s t r t s r t s r s r s s r tr t s s s s s s s r 2 s 2 2 s 4, , , , , , , , , , r r s r s r > 2 s > s > 2 ss s s r r s r tr s 1 t s rr s t s s t p < 0, 005 s rr t rr r èr tr r r ss t s r s s r é s tr s t s r rès r2 t r t2 s t t P r q t s q tr rs rr s t s r r 1 s s ts 1 t r s ss r t r rq é r ss r r ss r t2 s é t r r rt à 3 rés é str t st st ss r r è è r è s tr t s r t s à s r t r s rt t 3 s t s ér é s st té r ss t à s t r st à sé tr ss q t s r é s séq s ù s ts t s q ré é r q st ré s t 2 r s s s t s ér é s r r rt 1 t s str s s rés t s s t s s ê 3 t t s 2 é r 2 r s s rt t é à r r t t t à r st té é r q st t t q st s r é s s s s t ö t ött r r t ö t ött r r r2 t stré r q tr è s rt rs r r t ré r2 t
66 2 r s séq rsq s é 3 str rs q s s r s q r 1 r ss t s ê t s t 1 q r tr s r r t 2 t ès st r r r ss s é s t st à r ré é t s r t r t s r2 t q s ré 1 st t s s t r s t r tt 2 t ès ré t q s 3 str 1 st s tr t s é t2 q s rt s s à sé t st s t é s à éq r é r q s è t à éq r t t sé t ér q é à r é s é é ét q s r t s r2 t q s s t s s t s 1 r é s s é t2 à s t t q t r s s s rs s t r t s é é ét q s tr t s r t s é t s t rs s r t s r ss s sé t t s rt r t s t rs r à 1 r ss t t rt r t é ét q r2 t q ré t t t r é t2 q s tr s r 1 r ss tt 1 t t q s s st ré t q st s t t s tr s tté r t t r rs q st s r é r s 3 s str s r é è été ré é t é r t r s é t2 s rt rs r 1 r r s s s st r s 3 r s s ù s é t2 s t2 q s r ss t rsq s s s t és à s t ér t r s t s ss t r ss s é s t été s r é é t2 s rt rs s été tré à tt 2 t ès st sé s r s s t êtr très r t q t à s rt s r s s té ér r r é s t s tt 2 t ès ét t s r 1 q r s tr s 2 r s séq t r 1 r ss à é é t é s t ét t tr s s 1 té ér t r t r t tr t é r q t r s s t é tr s st t s s r s é s é s t st r tt 2 t ès s ét s r ér ts tr ts é t2 q s s s tré s s r étr q s 2 t s s s s ét té ér r s tr ts tr t s ér é s t t s 3 str s s é s ê s s r r é tr s r s s é s tr s r t q tt r t q r t r é r 3 st r s s s ès t 1 s é r q st r st s q s s s t 2 t t s t q
67 r ttr t s r r 1 r ss s s t s ér é s s s s r r r r èr tr tt 2 t ès s r t s r s r s s êtr q t s t sq st ér t st r s s è q s é r 2 t ès s t q rés t t ér t r t s s st t s r r r é s 1 r ss é s s r tr s è à s ré t t é s t t é r q r ét r r rt tt 2 t ès t s t s s t q s séq ç r ét r 1 r ss 3 s t s s s s tt s rt s s t sé s s t s séq ç t é t rès s s ér s 1 r ss s r é s s s r s à 1 r r s ér t s 1 r ss s t s r t q s s s q s s tr s s t s séq ç s t r s s ér r t t s é à 2 r t s r s s rt t s s s tr s è t r é r s s r r s r s tr s r ts q s s t r t t r tt t r r q t t s s s s t r tt s à t 3 str 2 tt t t ér é q s s ré é s 3 r é ô r étr t t r q t st r rés té r s é ts st à r ss s s ér t s t r tt s rt t ès s st s r t s r ét s r s r t t s s t s 1 ts t t r ss r s r r t t r t r r r t s s t s térêt r q t é s q r s s t è s r s s é é ét r 1 r ss s s t s à s s r r r ss r t r t s r r ss r r t r st r sé r 1é q tr té rt r rq t s s s rt r r s tr s r ts séq és s r r t s s r é st r s r 1 r sq s t s t à rés t t s r é s s P rq tt str té é st r st 1

68 % de transcrits Dmel genes Dsim genes No matches r P r t tr s r ts rt r és s r st r s r s s t s r s 1 é s r s q tr t s r rq q st r t t r tr t tr s r ts rt r és s r st r tr t s r s s tr t tr s r ts rt r és séq é té t ss é q é s s rt r s r é r r té r t t r s r t s t s r s s t s q é s s t rt s è s tr r ts s t s à rt r r s r é st r t st rq s s s t r 1 è rt r s r é s s r 1 t è s r sq s s s t s é s tr s r t st ss à ét r r à s tt str té rt r s tr s r ts r s r é st r s s r s s t s tr s r ts t été ss és à t ts è s ér ts rès rr t r s s P r 1 2 rt r à rt r s q r à s r è s t t s s r 2 s r tr r q é t r rt séq s q t é s r st r s r s s t s r s 1 s è s tt str té rt r sé s rs r è s r r r è r s è s t rt s r ts é q s t êtr rt r és s r é st r t tr s s r ts r r rt é ré ér t été rt r és q s r è s s s s s
69 Effectifs Effectifs Taux de divergence Taux de divergence r str t t 1 r s è s tés rt s tr st r t s s s r 2 s t r t str t t 1 r r s è s é t r t ss és st r t s s r rq q str t t 1 r s è s rt s st 1 è rr s t à str t r è s ss és é t r t s é s 1 tré s rr s t ê è P r rr r s s t sé s t t s rt r r 2 s t t s r t s é s s s s s r t rç s q s t t s rt ét t r t t r 1 t s s s r é é à ér t s é s rt t s2sté t q t s è s tés rt s tr s 1 s è s t t r t 1 r s r ts s r t s s r é é ê s t è s st r t s s ss és é t r t r tr str t t 1 r tr è s rt s t rr t à t tr è s ss és é t r t à r t r rq q str t t 1 r r s è s tés rt st rt str t rr s à r s rt s s st à r r q st ss r 1 t str t r s è s é t r s s t r t è s à t t rt rr é q 2 t r t ss r 1 q q s è s rt s très r ts tr s 1 s è s s s s té ér r rt s è s s s r ts s s r s s r tr r r és r s è s à ér r r s è s r s ér r à q rr s à str t r r s è s ss és é t r t P r s è s s s é r é rt r r é r r ts t ré r q t t
70 Effectifs Taux de divergence r str t t 1 r s è s rt s rès rr t s r s r t té s r rt rr é r tr str t t 1 r s è s rt s rès rr t 2 s r t s ss râ à r s s s t r ss é s è s 2 t été rt r és s r s 1 é s s tré à q s s ttr é ér t t è 3 s è s t r s s s s ssés tré s à è s st r t q r è rs st r rt s è s q s t s è s s tr r ts s s s s q r è st rès rr t r s r è sé r tt r st q s st s è s ér t t 1 r és ét t st t és é t ts st r t s s s s à ss é r t s r t r r tr r è st r rr s t à q t t s s t s s s s t rs r tr r éq t st r t r st è s t t s s s s st s è s ér t t 1 r és P r s è s 2 à s s è s à é t r r sq s ss t rt s été ss rès rr t t s è s 2 t s rt s 3 st r st à r s è s t t t s é q s
71 r è s ér t t 1 r és tr q r s 1 à 1 s t s t s q é s > < P P P t r rt 1é q t r 1é q 2 tt 2 tt P P t r t r 2 tt r s s s t r r t tr s r èr s s 1 t rs r ts tr s s t s t r s rt rés t s è s s tr s r s s s 1 t s é é s s r 1é q s r t r s rés t s è s s tr s q tr r s s q t té t t r tr 2 tt t é t s s t s é s s è s é r ts 3 s s t été r tr és 3 st r t t été r tr és 3 s s s è s ss è s s s rt 3 st r tr è s s s rt s 3 st r s r s s è s t s êtr s s s 2s s t s q st s rt r t rt t sq s s t é ér t s s t t é à t rô s s s 2s s s t s r s té s è s r r 3 st r rs q s t t rt s t t s 3 s s tt rt r 1 t s été é é q à tr 2s t s s s rt r é q t s r s s s r s s é r é t r t s t q s r t ê és r s 2s s rt t s s r s t r s t ss é s s ét t r s r s é t s r è s r 2s s s t s é q s q t ré s r s s r st r s t é t s t sq r s t s è s q s s r s st st r t s s tt 2s s s s r s r t s2 té tr s 1 s è s 3 t s s s t rt r é s tr s r ts s r st r s r s r r t s é t r rté r 1 è ét s s ss é s tr s r ts à è s ér ts q r s t s t r r è s ér t t 1 r és st 1trê t r
72 sq r r s tr t à tr t s t t 1 r s s s t r r t st à r s r s s q t s t s t s ù r ss r t t s t ér t s ê s è s s r s s 1 r és s t s r é s q r s P r r s t s s r 1é q s t 1 ès è s s r 1 r és à t r r r rt à 2 tt t q r s r s s r ss r s t st à 1 ès è s s r 1 r és s r t r s tr s s 1 q t r s r s t s t t t t q s 1 r s r s t s r s r s t s t t à r t s s r é s è s s 1 q tr st s è s ér t t 1 r és r s q tr r s s ss s tr ôté t t r tr t 2 tt t é t é s s tr é è s s r 1 r és à t r r r rt à 2 tt t s s 1 r és à t r r r rt à 2 tt s s 2sé s s t s r s q s s è s r s è s s t é ér t r r és 2s q t s r s r s s r r t sq à q è s t t 1 q r tt s r t t ès s 1 s s s è s 2s q t r s s t ré és s è s q és s s r ss s s r s t é ér t tés 1 ê s é è s s t r s 2s q t r s st r t é q s è s s t ss s t s t s t s r s 3 2t r s P s r s tr 3 s r 1 r és à t r s r r s r t r s 1 st à t r q tr s è s ê t r s rt r s s êtr ér t t 1 r és s s è s ér t t 1 r és t 2 s t r r és q tr è s ér t t 1 r és t 2 r s r s 2 t ès q st ér t st r é t 2 r s s s t é s 2s s r rés t t t r s t s r s st s è s t s t ss t rr 3 t t r st s r r rés té s st è s s s 1 r és à t r r r rt à 2 tt rs q r 1 t r s s t s r r rés tés s st è s s r 1 r és à t r r r rt à 2 tt r t 2s ét é s è s rr s t à s t r s ré è ss t t 1 s
73 Chromosomes X 2R 2L 3R Cytochrome P450 Métabolisme des cytochromes Glutathion transferase Glucoronyl transferase Turandot Autre Inconnu 3L 0.0e e e e e e+07 Position (bp) r s t r s q s è s s r 1 r és à t r r r rt à 2 tt t s r r ts 2s q s s è s ér t t 1 r és
74 r s t s r r rés tés r s è s s r 1 r és à t r r r rt à 2 tt P r tr rr r t t s t t t tr 2rr r t t tr s r s t t r t s t t r s s r r t tr s r s t t2 2 r r2 r s r r t s r t r t t r t t 2t t t2 r è s tés t r s r q êt r è s tés t r s é P s t té s r r rés t t t r s r q êt r r rt é é ss t rr 3 t t r r s t t t t t t r t è rr s t è s t s rs r rés t ts s t s r 1 r és s t étr r r rt à 2 tt r èr st s 2t r s P t s st s t t tr s ér s s r rés t s è s s r 1 r és à t r r r rt à 2 tt s r t s 1 r ss 2 s AG+PG 2 / AM+BM 2 tr s 1 t s r rq rt rés s 1 s è s t és ré é t s 1 s s t s ré é é s 3 st r rs ét s s r t t 3 é t t s P 2s s q s t r s â s tt r t s s r t t 2t r P 2 st s è s r 1 r é s t t à s s s 1 ét s t s ôtr rés t t s é s 1 t s s ét s t 2 r s rs r s s 1 q t s ér s s r é s tr s 1 s è s r st ss q s r s ét s t s r ér r s rs s r t té s s rs s s s t st r q st r t s s s s t té s ér t à r r t r t s st à r q tt t t t s r é s ê s è s st é t ss q s tr s 1 r ss t été s r s rès s s s st é é s t s s s st r q s s s s ét s r t s ét ts ér ts s s è s s à rs
75 Ratio d'expression Gotheron / Mayotte Cytochrome P450 Métabolisme des cytochromes Glutathion transferase Glucoronosyl transferase Turandot Autre Inconnu Cyp12d1 d Cyp6g2 TotC Cyp6g1 Cyp12d1 p Cpr92F Cyp6w1 GstD5 Cyp6a2 Cyp6a8 Ugt86Di Dsim\GD19446 CG2065 Dsim\GD21937 GstD2 TotZ CG10991 Dsim\GD17266 CG11659 Dsim\GD22397 olf413 CG13658 CG4500 Cyp6a17 Cad89D Dsim\GD15357 CG18528 Cyp12c1 CG13659 Dsim\GD15375 GstE1 GstD7 Cyp6a20 CG15068 Dsim\GD25680 Dsim\GD11688 CG6908 mt:trna:i Dsim\GD20102 CG3290 Obp56e Rab23 Cyp28d1 Ugt86Dd CG15661 Dsim\GD19889 CG31272 Epac Dsim\GD19273 CG7139 CG33301 CG16836 Dh31 Cyp6a21 CG6776 GstD3 CG9813 CG1319 CG18522 CG7194 Cyp6a23 GstE5 GstE3 CG13905 Eip55E MRP r rts 1 r ss t r s r 2 tt r s è s s 1 r és à t r rt r rés t t s 2t r P t s t t tr s ér s s st r s é t s st é t ss q s t t r t s q s s ré é s t s s s ré é 3 r t êtr s é s st r t sé s r t s 3 s s à s r ss s sé t ér t s 3 r r 1 1 r ss ét t rt t é t r t r t rs à s ér s tr 1 r ss s t q s s è s s t s s s à é t tr q t é r rs t s s s r t s ér t s s t r s r ss s st st q s ér t t rés é é r s è s ér t t 1 r és s 1 s rt t q é s s s ét 1 t s r t s été s r r t ré é r é t s è s s stér s s tt st rt t q é s ét 1 t s st s r s rés 3 st q 1 s s sér s t s t s q s t t s st t t s r st t t s t s r t t é à str t r é ét q s t à st q t ê à s é è s ré t r é é ét q 1 r ss t é t s s rs t s r r r 2 t P rt t stér s st ér t t 1 r é tr t s s tr ét t é t s s rs q s tr r s s ô s t t tr s ér s s s t t r P r s è s s r 1 r és à t r r r rt à 2 tt s tr t t è s s t t tr s ér s s s q tr s è s très r s r s2 tr s ér s s s è s t rô s ét 1 t s 1é t q s 3 st r
76 s t t tr s ér s s st sé t tr s s è s t à è s t t 3 s s è s t s s è s s t sés ss s t séq t rs r tér st q s q s t 2 t t P r s ss s s δ t ε s t s é q s s s t s t s t s 1 s r s r é r t t st r ssè s s é δ t q t r3 ε s t s s s s s t rt t é é s é t 3 st r t 3 st q s q s èr rô s 32 s s t t r s s è s à r t t t s s s è s s s r t ét s s q s s t s ét s r t tt 2 t ès st ér t s s r t s tr t étr t à r t s t r sé 3 r 1 sé à s st s tr rt s s r 1 r ss s s è s t t t q tr δ s t tr s ε s r èr ét t ω tt ér 1 r ss s é à t t à r t t t té à ét 1 r s st s r és rt s s r t s rr t r été sé s r s r ss s st st q s s t s r t s à s s r è s t s 1 r ss s è s s t t rt t 2 t ès t t 2t r s P P r s è s s r 1 r és à t r r r rt à 2 tt tr é t tr 3 2t r s P s q tr s è s t t s s r té q s r s s è s rés t t s r rts 1 r ss r s s rq és r r s 1 è s s r t s 1 r ss s s é és tr s s s t 2t r s P s r 1 r ss s t r ç s q 8 à 16 r s s t è s r r 2 r 2t r s P t s 3 s s ss st s t t r à très rt s r 1 r ss st t t tt è s q r 1 rô s P r s rô s 2 ét 1 t s 1é t q s t s 2t r s P s r 1 r és s t r rt s r êtr s é q t q és s rô r t r t 2 r s t t s t t tr s ér s s s q t t s t s s s à r t r étr t s t ssé ss t t r t t à 1 s t à s r ts t sés r r t r st à r s st s tt t t st st t t t s s êtr st é r
77 Pas d insertion Insertion d un élément Accord D. melanogaster Duplication du gène Insertion d un élément HMS-Beagle Insertion d un élément P Élément P tronqué D. simulans Pas d insertion Insertion d un élément Doc Insertion d un élément Juan r é t t s è s 2 s 3 st r t t t s s s rt é é t s t r été é r t tr ét r s tr r q tr s t s ér é s ssè t s s r t s ér t s sq tr t t r tr s rt é é t tr s s à t r s s s s rt st r rq 1 r é t r t 3 q sq ê s â s é és s r 1é q t 2 t s été 1 sés 1 st s tr t s s r 1 r ss s è s s s s è s s t ts t q tt t st tr s s à s r ss s é é ét q s 2 st r é t P r s è s s 2t r s P s t t s r 1 r és à t r r r rt à 2 tt s tr è 2 è sé s r r s r t r s st 1 r é r 1 s s à t r q à 2 tt r r è tr é t s ér s 1 r ss tr s t s r é t r st r st 1 r é r s s 3 s â s r é s q 3 s â s r s tt r t t tr s s s q r s s r t st
78 s è q st s r 1 r é r r r rt à r q s s 1 ét s s q s ôtr è st r tér ss t r t t tré q s s r 1 r ss ét t é ss r t s s t r r ttr à st r rés st r s t s t t r t r s s ss à tr s s t s r t 2r t r r s rt s t s t r s st r s r 1 r ss st à s rt é é t tr s s r t 2 r t rt t r st t s ét s s ssé s t s t tré 1 st s s 1 è s ér ts t è 2 s s s rt t s r s à è 2 q é t s s rt s s q r ré t s ér ts è s s à r 3 st r t s s 3 st r s s 1 è s s t rré és à 1 r ss s s rt r t à s r r r t t r r été ét r s ét s é s s t t t à rés st 1 s t s t sté s é ér t s r s s rq é s t t t rt t r st t t t 1 r ss st r str t à q q s t ss s 3 st r s t s t st 2 æ str q t r s r s rt t r st t t 3 s s t t tré r rq r é t é r t é t s rt é é t tr s s t 2 s t t r r é é t ér t s r é 3 st r r tr r r t r tr t s s rt rs q è s rt é é t tr s s st rés t r à réq s r 1 r ss è à t r s s r r é 1 st s rt s s 1 t s étr t t r s s s ét té s rt s s s s é s rt tr é é t tr s s à t r s s s é rt t s t s rt r P r st r s réq s è q s s rt s s 1 t s s s é t2 é s ss s é s s s ér t s r q t s t t r s s tr é s 32 t s r s rt t q tr étér 32 t s s t réq s rt st é tr t t r à st é à rt r ér t s ét s s t r s r s t r s s s r é s 32 t s s s s rt t 1 s étér 32 t s s s s rt s t réq
79 s rt st é tr t t r à t t s ré é é s rt s r t r n = 10 t 2 r s rs r s s à t r é t s r s ét t r s rt r n = 10 2 r sq s ét t r s rt rés t à t r rés t è r q st 1trê t ré t r str t à rt s s s t s r ét r r t q t s rt é é t t s s t ssés à ôté tr s rt t t é q s t séq é s é s st r q s s t ssés à ôté s rt t r t q s t r rs s 1 r èr s 2 t ès s r rq r é t tr st r t s s s ss à tér r s 1 s è s rré t rés st 1 s t s ét 3 st r tr rsé 3 s s st r q s s s s rés s t s rt é é t tr s s s é t2 s rt t sé t és st r é r s ét s 2 r s r t s s rt s t t tré 3 s s très rt ss 2 r s s ét t sq à s s t r é é t t t t t tré 3 st r ê tr s ét t sq à s s t r s rt s ss s rt t s 2 r s s t s rq s 2 sé t t r t t s s é t t sé s réq s r t st r s s t s ét t à éq r r 2 r à s rt sé t rr t tr r s t s t r q r é rt à t éq r s s s sé t r t t é r q t s é rt r éq r r 2 r t ss rt s t s s s û st r s s t st χ 2 r s t rès s t sts s t s s é rt t s s t t éq r r 2 r = 0,33 r t 2 tt t 0,39 r t t r P rq s tr s sé t é r t s s s t s ér é s t t t sé t t ss t t s r s 32 t s s s s rt 1 ét t t t t t ç très r r s s t étr t sé t s t s r s réq s é q s t sé t s st s 1 r é s r s s s é é s r t r s q s r r
80 s st q q s réq s é q s st é s s r s â s tr t s é rt à éq r r 2 r t é t s t rr r s r réq s rt s t r t t t t éré r s rt s s t s ér é s st r t t r r té tt s rt r q t êtr s r r t t t q rt t t tré rt t r r t r s s st r rt s s è s rés st P s rs 2 t ès s t 1 q r s s r t s r st ss q s rt s t r ré t s t étr s tr t ér é t q s rés à 2 tt s 1 q r rt ré t r r t s é s ér s à 2 tt s t ss t t r étr t s î s r s s r t s ét t s s r tr r à 2 tt è rés t étr s 2 t ès st tr sé t s rt s s t s r s t t 3 s â s t s s 1 t rés s rt r q t t r sé t 3 s s 2 r s s st rt t r st t 2 ût r 1 r r t st t êtr é 1 â s t t t ét é s s ts r és t ss 3 s â s t t tré s ts très r s t é t é ét q s q è rés st st tr t s t s tr r ût st ss é s s t rt é st s rt t r t tr s s é ût t t st très r t t r s r q r t réq s rt s r t é q éq t r t è s q ré 1 str ss rt t s r 1 r és à t r r r rt à 2 tt s tr t 1 è s rt t à s r t t t t tt st sé t è s 3 st r r r r é r t été t è r t r r ré s t t 1 s r q és r t s t ér t r s tr r t 1 t Pr t s P s è s ré t str ss rès t s t r r t ts t s s é 2 r t t r s é s s ré t s tr t 3 st r q s è s r t t t t é t êtr t és r tr s st r 1 s st s r rés r t
81 s ét s t tré ré s t rs st t s t s tr r s tr s s r 1 r ss r t st t t t t t s t t r ré à t r s s r r t t 1 r ss s tt è s t s ér s t êtr s t st s t q s st à r séq é s r r s t s t rés 1 è s r s r t t t s r t tér ss t s r r t è 3 s s r r q st 3 st r ts r ss r s séq s r s s s é é s 1 t s à s s r r r r t r r 2 tt t s r r s t st r sé 1é q q stér é rt t êtr t é r 1 r r r r té r s r s s ê s t ét t s r r s ts 1 r ss és t r ss r str ss ét s r r t s r s s s t t à t r r ss r t r s s t t s r é é r à é s r s r s s s r 1é q r r rt 1 r s s s r t r s s s s ré é ré sé t sq s s s s r t st r r st s r s s t t r t r é rét t s r s s s r q st s r r t r é t s r 1é q s r t t s r s à 1é q t té s s ré t 2 tt é é ôté s r 1é q t tr s r s r ss r s t r s î s s s r é è s ér t t 1 r és tr s 1 t s t P r s è s s s s t s 1 r és s r 1é q s tr s ét t s r 1 r és s r t r t st s r r rés té r s è s s r 1 r és s r 1é q s tt st 2 1 è s s r s t ss t s è s t rô s é t r r s s s t ss 1 r és s t st 2 s t s s t s t s s t2 2 tr2 s q é s s st r s r 1 r ss st t êtr tr st té rs t rr t r s 1 è s s t s
82 r és r tr s t s ss q s s r té s s é s t r s s t s ts r s t s r r t s tr s è s t s t s très rs s t é r t s 1é q à t s s r r s s s ê t è s s t s 1 r és s r q s r 1é q st s t t s q r è s s r 1 r és s r 1é q t st s r 1 t = 2, P rq tr 1é q s s str ss t r s r s s r r q rt t s2stè t r st r q s r r è s s t ér t t 1 r és t tr s t r s t s s r r rés tés s st è s s r 1 r és s r r r rt à 1é q s s t r s s t és à é s t r s s 1 é s ét s è s és és r s t r s té r s r rés té st è s t s P t s t r s P ér t s t rs té 1é q st stér q st s s tr r t r s r r s s t t tér s q q s st s s r r t q t rt t t s2stè t r s r s s s s s s tér ssés 1 r tér st q s tt t s r s s t ré s t r é très r st s r s t r s q t tés P t t 1 s r s ré t s s t r é tr s s t s tér s r s t t s s t 2 é r s t t t t 2 r q t 1 r ss t té s s s r r t t t r é tr s tér s r é t r r t t r rr2 t r tr t r t r 1 tr s s s t r é s r ré s t r ré s s t r ér s t r t s s t s s r s rs t t rs r s r t q ré t t s è s s r ts tt r èr st r ss 3 s s q t s rt t rô t r tr t t t P t été t és à r r t st r é s t rs s tér s r é t t r tt r s r st r t t s t rs s r s t s s t rs s s r s 2 t t tr
83 r s t è s s r r rés tés r s è s s r 1 r és s r r r rt à 1é q P r r s s t t r t t r r r s s s r s s s r s s t t r r s s t t r r s r s s t t st s r s s s2st r ss t r r r s s r r s s t r s r ss s r s s t r t t r r s s t str ss tr r r tr r s s r s s t r s t t r tr r r rt r s s t st s s r s s t s r s s t s t2 r 2 s t t t 2 t r ss s 2 t r ss t 2 t r ss t t r s s r è s tés t r s r q êt r è s tés t r s é P s r r rés t t t r s r q êt r r rt é é ss t rr 3 t t r r s t t t t t t r t è rr s t s s t r s és t s è s q és s s r ss s t r s
84 t r tt t è s s s tré q s t s r 1 r és 3 s r s s é é s s r r r rt à s é é s s r 1é q s 1 s r 1 r ss s s t 1trê t r s t r rt r t r à r rt r t s t s é ét t s tér s t s rr t s s s t 1 q r tt st t st à t r q rés r r s s s té rt té s â s t s t s rs ss t t sés s tr r t tt rt té 2 t été 1trê t s t t s s t s s s à 1 â s r r t t s2stè t r s r ré t 1 s t t s r t s s st t t t s 1 r q rés r r s s t r q é rt té r r t êtr t é r é q s r s s s s rés st t s 1 r r s s s ét s s r s2stè t r 3 r s t r sé s r s t s té r s s t s t s t q 1 s t t s s t s s s t s s t s és s t tr t t s s tr ss tr ét st r èr à é r r t s2stè t r s r 1 s t r é s t r 1 t è s t êtr tér ss t r s r r t s r ss s t r s t r é t rs t t 2 t r è r r r ré t r r s é s à ttér t r s s s s q s r r s s t à q tr t t st é s r s s s t r t été 1 sé s à s t s rs é s s t tér s r s t s t é t s s t s t t r t s à q s t r tr r s t r t s r t s t r s tr s à s r s r tèr t t r s rs té P r 2s ét é ré s s s t s r t t r tér s r s r r s s rés ts r 1 r ét ét é q q ét t s t tt ét é s s s ré 1 r ss s è s s2stè t r s r és s tr ét s tr s 1 r ss é r ts s ttér t r s s ré s 1 t s P s s q 1 è s q és s r ss s r s tér s P t 2 t Pr t P P t s r é s ét r r t q t t s è s t t2 t t t s s P Ps t rô t
85 r r rt 1 P sq s s t q és s r ss s r s tér s t st t s t s s ré t t à ré s t r r tr t t s P s t s 1 P Ps rès t tér q rès ét r r t t s tr ét tt t s s s ré t s tr ét t s s s st r rr r s r s r t s t s rs s t r é t r 1 t s sq sé s s r s s r s r t é s r s r s s t s r tt t r r r r r t ré s t s s ré s r r r t rt t q t à s 1 à q ré ts rt s s è s tr t t s r à r q é r qûr s t q t t s tt s P P t t êtr t r tr s t ré rs st t q t t t r st r s t P P q tr t t r t t té 1 r 4 1 tr s è s r s t s t s ts s tr ét q s r r t s rés t ts tr stés s t t êtr s 1 tés 1 s t 1 r r s s s tr ét à s rs é s t s t s tr t s r s 1é q à ût r r r t è s ét t s r 1 r és s r 1é q r r rt à rs q è s ét t s r 1 r és s r r r rt à 1é q s s é t 1 é s s r r rés t t s t s è s s s st s r s t t r s r s t r t s r 1é q t rés t s t r s t s s r r rés tés r s è s s 1 r és 3 s s é é s s r 1é q r r rt à s é é s s r 2s ét é s t r s tr q s és t 1 r s è s s è s és à r r t t s s s è s s 2t r s P r r r st sé t è s 2 t t s r r t s t s t rs r Pr t s s r té s s s ss r s P s r té s sé P r t Pr t t è t é r s t 1 tr s Ps q s t r 1 t s té s q é s s r r t s q tr t r t ê tr 1 r ss q s t tr s tr st r tér st q r s è s 1 r ss é q st
86 Attacin A Attacin B Attacin C Diptericin Rapport d'expression h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A Rapport d'expression Diptericin B Drosocin Cecropin B Cecropin C 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A Listericin Metchnikowin Drosomycin Defensin Rapport d'expression h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A 1.5h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A PGRP SB1 PGRP SC2 Rapport d'expression h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A h 3h 6h 12h 24h 48h 12h 24h 48h 96h B/A Blessure infectieuse, 1h30, 3h, 6h, 12h, 24h, 48h Infection fongique, 12h, 24h, 48h, 96h Banane / Axenique r t s t s t r s s r r r rt à 1é q s ss rès t tér q r r rt trô é s ss s r r t
87 r s t s è s s r r rés tés s st è s s r 1 r és s r 1é q r r rt à P r t r r s r r t r r t tr r s r s s r r t tr r r rt t tr 2rr r è s tés t r s r q êt r è s tés t r s é P s t té s r r rés t t t r s r q êt r r rt é é ss t rr 3 t t r r s t t t t t t r t è rr s t r tr s t s 2 tt 1é q 2 tt t 1é q t r s t r s s é é s s r q s èr ût r r t r é s r ût st r t t û é sq ê t é é s r 1é q rés t s s r tér st q s P rq s r s s t êtr rs s st ss q s s ré té s s r t rr r s r r q r t r é q st à r tr té t été t é s r s st s s r s t rs rés st 1 st s st ût s r s r s s q è r t rs à ss r r t 3 s s 1 s t 1 st s t t r q té s r r té t tr t s r t 3 ï s r s r s s t r 2 t t r t 3 st r s rt s è s Ps tr t t 1 r ss s t à 1 s t 1 st s t t s s ss r s â s s t rs é r sé s t t r t rés t t r rs s s r t t é t 3 1 rés s t êtr à r ût r r t tt 2 t ès s r s tr r r s è s s r 1 r és s r s è s ét s ét r r s è s s 1 r és s r q s r 1é q r tr s s è s s s q és s ét s ét t t s s t té 3 s s st s q 3 st r ré t rés st r à ét t q t s ss t r t r s t t 1 tr s è s s t s r 1 r és q s t t tt 2 t ès str 2
88 è ré s à ét t q st té è ré s à 2 1 t q ssè à t êtr sé r s r t r t ê s s s s t s r ét q s r t r st r t r t t té tr s è s ér t t 1 r és s tr ét t été s és r ré é ts tr 1 q és s rés st s s té à ét 2 s s 1 r és s r t r t t s r 1 r és s r r 3 t t ét s st tér ssé à t r t t s r r r t 3 r s ût r r r t t ss êtr sé r tr s é è s 2s q s és à s ér s s st tr t s r t s é ér t tr ts rs t 1é q à r è s és P r s è s s 1 r és s r q s r 1é q tr é t r è è s s s q és s t r t s s s s s t tr t t é tr r tr st s ré é é r 2s s t r s t t s 2s ét é s è s rés ts s r s ttr é t r r s s ér t s ré t r t t rô rt s s è s s t s r st r t t r tr s s t è s rt t é tr s st é t ttr r r s r 1 r ss à t rt èr tr s t s t è té t s 2 s t s s s ét s t r è s t t t r és s ssés à t r tt r t t tré 3 st r s r 1 r ss è s és à s t r 3 t r r r rt à t r ç s s t é s 2 t ès q st û à ér t s s r r rt à t r s s r é tr s t s 3 t ss r s s t s t rès éq t r s r s s t t t s r r rt à t r r s P tt t s r 1 r ss s è s rr t êtr ss é à réq s r s tt ts tt r t s s s é s s r t s s s s t r ss r s é s tr t t s t t r r tèr 3 s s 2 t st r s r tt 2 t ès
89 s ss é ér tt ét s st é t t t s r é t s t s t r s t s t s s t t s à r r t t t s s t r t s tr s r s s s s é s2stè s 1 s s r ss s s t q és s é t s t s t s s è s st rô 1 r ss s tt é t s s r st r st ét é s t s s tr r t r t é r q é q t st r q s q t é ét q s t s s s r s s s t s r t r t r t à s ét s é q s t é ét q s tr ét s s r s r tér s r r 1 r ss tr s è s t tr 2 r s t r s é q s 1 r à s rô 1 r ss s ér t s s è s t s rt r t s 1 r ss 3 s 2 r s q rr t êtr st t t tés s é 2 r s r s é t r r 1 r ss tr t s s s t s r t ss 3 s r r t ér t très tr t s s ssé à t r s r s t s rés t ts s r s s s t sé r séq ç t é t q r t rt rt r s tr s r ts s s s s tés à t 3 str t t ér é ttr é s r ss s tt t r t t s t s rs s t r t s s t r s s r r t s ér t 1 r ss tr t s s s s r r s s t s r s 2t r s P t s rô r s t t à r t P s s s t r s s s é t 1 r ss sé t rs s tr té s é q r s s ér t s t q s ét 1 r ss é q t s t r r s s r 1 s ts tt t ès
90 ér t 1 r ss tr t s s s ér t r s tr s 1 ét s s s ré é é è s ér t t 1 r és tr t s r ç s t r ét t é t t s t t ét s t r r t t s s è s ér t t 1 r és tr t r ç s t s t r P rq s ér s tr s 1 ét s q t t t s 1 t sé s ss â s 1tr t r s t r â s â s ét t é èr t ér t s t s t rs s ét r s q s ét t é t s 2 s r s q t s ér s tr t s t tr ét s s 1 r q rt é r q rt t r è s ér t t 1 r és s s r s t é é t r s t P r s è s ér t t 1 r és tr s t s s ét séq ç s t ét t r rés tés s r ét s ré è t t ê s t t s è s ér t t 1 r és s s r s s t st s r 1 t = 2, s t 1 1 s t s ét t 1é à s ét s tr s ét séq ç st q t 1 q r s ér s s 2 t ès s s t t s q t t r s ss ér r à r séq ç r t t t t s t t r r t r s r s séq rs t é t ét st t st q t sé r 2s s é s séq ç s s st é r q q ré r t s 1 s t s t s s t sé r s r t st t r q s r s s t s tt 2 t ès st r s s r t é s s r t r t s rs é ér t s s s s s ré t s ér s tr s t s s s ét és r s t ét s r st r t s t s é s s r é s s rs é s r t r ré é é 1 ér ss t r s r è s ér t t 1 r és tr t r t ér é t tt r t r t P t êtr s s ré t s s
91 s r s ér t st 1 r ss s s t s é s s ê r t tr r t s ér s q 3 st r r q sé r t st s s tr s 2 t ès s st ré t t t s s s tr 1 t à s s r t s s t é t q s rs 2 t ès s s t q é s 1 è 2 t ès ét t ré s r t rs t à s ss t r s t r s r s t s ré t ér é s s 1 è s ét t ér t t 1 r és tr t s à s s tr ét s t s tr ét r séq ç s t 2 t 2 s 1 2t r s P s t q és s t t t s 2s r s s t s ss s s r s è s s rt s r s 2t r s 1 r ss t t t s ét s tr t ss t t t t s r ss s ét 1 t t t êtr ré t r st s é q r s ét é P t r tr r s tr s 3 tr s r s s s s é à q s r 1 r ss è s ét 1 t st tr é t ré é é 3 st r tt r t r t t s t rs t t tr s é è s 2s q s q és s ér t tr t s s s t s r é s r 1 r ss r è s q és s ét s s s r s è s r 1 r é à t r st t t t q é s ét s r s r s st r q rô r s ét 1 t é ér q 1 q tr s r é r rs s s r s s 1 r ss tr é s s s s t rés st t s P r t s t rs t é t s r é s r 1 r ss r q è s q és s r t s s s tr st t t t û à s t t t s s r r rt à t r s s r é r q q s r t rs sé r réq tt t s s s é é r s r s r 1 r ss s è s és à r t s s s tt r t 3 s s s ér s t t s s r s s s t s s r é s 2 t s ér s s r ss s ét 1 t t é t été ré é é s r s ét s 1 r ss s tr s s è s s r s tts t t rr r
92 tr t tt r èr ét ré é é t t t tr s ér s s s t à 1 s t à s s ét q s r t tr s s s s è s s s s s ér s r s è s q és s ét 1 t t é t été tré s s ss ç s s r r t t r st t t rs 1 q é t st r tts t t rr s tr s s t ss 3 ér ts 1 s r és s tr ét s r s r s str t s tt ét s t rs t s r é s tr s t s très rs r sq s r tr s t r s t s ér t r t s s r ss s ré t q t té r té s s r t s s ét s r té q s t s r é s é t s 1 ét s 3 r r t s t s t rs t ét é r 1 r ss tr q tr s ts r s ré é s s t r s s s s é à é q é s tr t rs rés t ts s èr t q s ér s ét s t s ré t r té q ss t êtr s à sé t s r s s ts t sés 3 tr t t é t s r é s t s ét s s r té s t t r é é è à t t à s t rs r t 1 P t êtr st é t r s é è 3 r 3 s r s s t str t t t à r t ss s t r t t 1 ts r trés t t t t t ér t r st tr r ètr r t r q s s è s s t 1 sé s ét s r s tré t t è s q és s r t r té q ré t à t ér t r s t s t é t ré é é t è s s r ré r str ss t r q s t s r t P s s é q é s tr t s t t tré q s t s s2 tr q s s s è s r s r s t t r t s r t s 1 r ss q s t s t r t s t s 1 r ts ér ts s t ré é é s ér s r s è s q és s tr s rt stér s q s tr s s ts ét s s t r sé q s ér s ét t s à t t t t à t ér t r s s r t s s t s r s è s é t tr 1 r ss é q r sq rs s ts 1 r ss s r t t 2 é s t 1 s tr s ét s t rs s r s t tré s ér s s è s ét 1 t t t t s 2t r s t t
93 és é s t s ss é s 1 q tr s 2t r s P 3 st r è s t s ss é s P é t r t t 2s P 1 ét 1 t s 1é t q s rés st 1 st s P t r s 1é t q s t ér s P 2 és st 1 s t s t s 2s q s r s 1 P st ss s s P t P 3 s rté rés 2 P st 2t r s t r 1 s tr s s s t r s 1 s é s tt t r t r 2 r s t s ét s tr t s tr s très rs t é t é s s è s s r t s q r tr s s rs ét s r t t t ét s s s r s ré t r té q t t à t ér t r t ét 1 t t t ss é ér t r t t à s 1 és à s ts ér ts s r t tér ss t 2s r r è 3 ér ts r s s s s s r ts s s t r sés ét r r à q t ré s à r t st s é q tr r à ér ts t 1 s s 2t r s P rt t é à t t ss t t rô s s 2t r s P st s s t é q s s s rt t s r è s t t s s s s t s è s t t s r t s s t é és s s é s r t s s ss s 2 r s rs rô s s t 1trê t rs t s2 t ès s 2sté r ï s t r é à ét 1 t s 1é t q s st s rt t r r rô q s és és s t ét r é é ét s t s r s r êtr t t à r 1 s ts q t t s t r s s à ét s t s é ts 3 s s t s tt r t è s s s q tr r 1 s 2 r s t rés r è s q s 3 r s s q s t s ss é s
94 s 2t r s ér t t 1 r és s tr r èr ét s tr s s t s rt t ss t t 1 s P t P r 1 rt èr t r s P t s t êtr ss t t q és s s ét 1 t s 1é t q s rés st s 1 st s t s s rt t s t t sq s t s P 2 ét t r rés tés t 1 P q t r3 P t s r rés t t r s s P t P t 2s r t é t à é t tré ér 1 r ss ss t t té à s s rs q s s P t êtr 2sés s ét r séq ç r s tr P P P t P s s s è s ér t t 1 r és à s s tr r s t r séq ç t s s r s t r ç s tr t r r s t 1 2t r s P 2 q rt t P t 2 q rt t P 2t r s è s ts t à 1 r ss é r r t ét ré s 1 1é t q s s s 3 st r tré s é té ré s s 2t r s P t tt ét t sé s s é é s t t tré s s r s tr s r s s è s ét 1 t s P s t t tr s ér s s t s stér s s s t rs t t sté t 1 1é t q s ér ts s2 tr é r t t r tr 3 t sés r t r s t t é t r s t tré q s tr s t s è s èr t à s s 1é t q t sé r t s ss s s 1 t q q s è s tr t tr r t t st t t s t é ér t é t ér t t 1 r és s s 1 ét s s tr s è s s t s t t ts 3 â s t s r s 1 1é t q s t s s tré r s tr s è s 1 r ss st t t s rt s t t r q r q t 2 ss s r 1 r ss st t t 3 s s r r rt à s ét s r tr s ér s 1 r ss s r és r t t r s è s s q q q s tr s q s t s à s 1 ét s t é r t t ré s s rt q tr 3 r s t s s r é s r t t 1 r ss ss s é s s r
95 s tr s r és q t t à tr r q t t 1 r ss s t s s tr s s r és rs 1 s t t à s 1é t q s r s s t è s st t t t s r 1 r és s tr ét tr s s t ts r tr 3 t t s s t ts à ér ts 1 r é r t t t tré r s t s été tr t r rs té r ts s st ss t r q s r ts s t ér ts 1 t stés s ét t s s é t r é à r r è s é s t s r s è s s r s s r s t rt s s è s 2 s t s r rés tés tr s t tr é s q t s s 2 tr s é s 1 t s st t r rt 2 s t r ç s r r rt 1 tr s t s r s s ér s tr s tr s t s sq t r ç s t t q à rs q r rt 1 r ss st r s t t t s t s s 2 s t 2 s s r tr ér tr s é s t 2t r s t ré s t t t t t t 1 r ss st t t q s s s r é 2t r s s t r t rs s P r s s t t s r s 1é t q s rs q t tré 1 r ss st t t s t s s é s q tr t rés st à s s st s r t t è s s t à s t s t tr t s è s rés st t 2 P r s t è s s 1 tr t t t s r 1 r ss à t r s tr s 2s s tr ét é t ré é é s 1 tr s 2t r s st t t t s r 1 r és s t r ç s s 1 sé st s q s èr t t s t s è s P t 2 r r tr t s r 1 r ss 3 rt s é s st r rés st t s P r t tt r èr ét é t ré é é è 2 q st é t t s é rés st t 3 st q t s t s s 1 1 è s tr ét t s r été ré é és sq à t t r ér t s r s s r s r s s ét q r s r 1 r ss st s é q s s ê t t r s ér s s r é s t été t s s t r à é ér t r t t tr s s s 1 s ts r s r ss s é é ét q s t s tr s ét s t ss t t rté s r s è è st r tr
96 Rapport d'expression Femelle, Phenobarbital Mâle, Phenobarbital Femelle, Atrazine Mâle, Atrazine G / M ~ ~ ~ ~ ~ ~ ~ ~ Cyp6a2 Cyp6a17 Cyp6w1 Cyp6a23 Cyp12d1 d Cyp6g1 GstD2 r t 1 r ss r 2t r s q és s s ét 1 t s r s â s t s s st és s t é r t s t à tr 3 r r t 1 r ss st t t s t t r r r rt à t 2 tt s s q t s r éré s r rr s t à s t s 1 r ss q s t s s t s r t st s s t t ér t P r q è s q tr r èr s é s s t t ré s t
97 ét tr à s q s è s rs q és s t t à rés st s t t q s tr st r t s s s q 2 é t tr s è s ts 3 s s q ér t r t t êtr êtr 1 és 3 st r t ét t q r s t s è s s s tré r s ér s tr é s rés st t s t é s trô s t t r s 1 è s s r és s tr 2s tt ét s rt t r é rô ré ér t 2 t t t r rs té s s str ts q è st ét s r s 1 è s ts tr ét t rr s r à s é té tr t étr t s t é t êtr s 1 t s s t r s tr 2s sq s r s s t s s é ér t èt r t r 1 r ss s 2t r s P été 1 é 3 r 1 r s s 3 s ét s t s ré r 1 r ss s t s très rs s t r à ré t r ss t r s ts 2s q s r ss ss 3 t 3 tr s r s s s ét s t tré s ré t s rs s à s t rs r t 1 t 2s q s s ré s à s 1 é s s r s 3 t été tré s r 2t r s P 3 P r t r2s s r r è s 2t r s P sq t r t s tr ét s t rs t tré q s 2t r s ér t t 1 r és ét t é ér t r r és 2s q t s r s r s s s 2t r s ré ss t 1 ts q s s t r t q t q r s t t s t sté st ré rt r ts str ss ts q s s t q s r t 1 t ér t r tr ts 2s q s t 1 3 t r s s t s ér ts str ss t r s t 2t r s P ér ts s r ét és rt s tr 1 ré ss t à t s str ss s 2t r s P tr t é t rs té 1 r ss t ss r rs 3 t t t tré q s è s t séq st r s 1 r t é ér t s s ê s t ss s tr t êtr û à ré t s r s è s t és r t s t s ss s t s 2t r s P rr t ê êtr t sé té t r t été s éré ét t é t r t s s st t
98 1 s 2t r s tr t t t tt st rt t q é s t t 1 1é t q s r trés r s t s rs très 1 2 t s s é à r é tr s è s tr t s 1 s t t r sé t è s t s r 1 r ss t rs t t ét é 1 r ss t str t r è t t é 2 s r s é s st r sé t é s r t r r r rés st t s é s trô s s é s rés st t s tr t 1 r ss té r r rt 1 trô s t rs t r t s t s é q q s t t s tr s 1 é s s s t rés st t t t t rés é é ts ré étés ss s é é t tr s s à 1tré té è 2 s s s s r q t st té t t tr s r t é rés st t tr è s rt r t r t t t s t é rs s é t rs t str t r s été r é s r tr s é s té s s t r 2t r 2 été r é à rés st r st t q t t r s s t s été t é st à ss s r 1 r ss q èr ré s st t3 t t t t tré s rt é é t tr s s s ré è s é rés st t s 2t r s t rt é à t t à s rs ét s st r s è r 1 ré t t ss ét s rs tr s t s t s ss rs rs 1 t s r s t t s st t s tr tr s r ts é q s è 2 tr ét tré t 2t r 2 s t t à t t t t rs 3 s s t t r t s t s s s s t rs tt ét tré s r ré è rt r s s t s s ss t ss t 1 stér é rt t s t t s t s t t s 3 r tt t s r q rô s 2t r s P s t t st s rrêt r 1 ét 1 t s s é é ts tr s s s t t q é é ts t è s és à t t s r t t s q rés é é t tr s s 2 s tr ét t s tr s s èr rô rt t s é é ts
99 tr s s s s r t r ts t t s s è s 2t r s t t ér é tt 2 t ès r r t s rés é é ts tr s s s à r 1 té tr 3 è s 2t r s 3 st r t rô s ét 1 t s 1é t q s t q q és s s2 t ès 2s s trô é t s t rs t s r é t s é è ts s rt é é ts tr s s s t r s t 2t r s s r s t q és s s ét 1 t s s t r s q 2t r s q és s tr s r ss s s é é ts s t é ér t sérés t è s t r s ét t r é t s t t s r t s r s r r 3 t s s r t s tt t t rô r s é é ts tr s s s t t q é é ts t è s 1 r ss és à t t t té s é é ts tr s s s st é ér t st é s str ss 2 t 1 s t à s 1é t q s 1 t r t s é é ts t r ttr t t t r ré t è s à ts rs s 2t r s P r s t t tr s ér s s s s r rés t t s é é ts tr s s s à r 1 té è s é rt t tr ts tr q r rô st tôt t t r r té à tr s s r r t ss t s st té 1 r ss r q s r s è s é ré t à str ss s r t s r r t s t è s rés st s r t t s t t s ré tr s r q é s ré ér t t r s é é ts tr s s s t s t s séq t tt r t ré s s s r s s s r t s é r t s ss s s r st t t s é t t r t r r té ré s 3á 3 t t 1 é s s rt s é é ts tr s s s ré ér és s r 2 s r r r r s s s rt t t s r s tr 3 s rt s q s t s r é s s q tr tr s r q t t s 1 r ss r s rt s t r îtr r t t s s t sé s r tèr s str ts sé t s r s s rt s r rs t t ré 1 r ss tr s t 3és s t tré r 1 r ss s rt r s è s q tr t s rt é é t tr s s s s 1 s è s t s t é t ré é é q s è s rt rs ê s rt é é t tr s s s s 1 é s s rt str t é t s r r 1 r ss s t q s rt st s s r 1 r ss s q s é é ts tr s s s t t à
100 s sér r à êtr r t s s s è s q r rs tr t s r r 1 r ss è s s tr ts t tr s ét s t s é r t s é é ts tr s s s s é t ré t r s r à té r r rs t P r r t t tré 3 s r rs q s é é ts tr s s s ét t r s s s r r 1 r ss tr s r s t r t r t t tré tr t rt t s é é ts tr s s s 1 séq s ré tr s 3 s séq s ré tr s s r t ss s s é é ts tr s s s s s 1 s st t é é ts tr s s s t s r t q s é é ts s t t t t s r s t s é q s t s rt st é ér t é étèr s r t t s s s r t s é s à t t à r t tr t q s tr s t t 1 r ss s t rs q q s s q ré ss t rt èr t t t s q é s s ét 1 t tr ss à q t s r ts t été tr s r és r êtr sq t2 t t q s s ét t r t t st t t à tr t s é s2stè s ét t ss s ts ét q s t t s s r s t s t q t t r r r s ét s q t 1 é ér t 1 r ss tr t s t r s st rs té s tr s s r és rs s q q s s s s t é s r t s r q rt é r 1 r ss st s t t s r é r é t tr ér é ét q 1 r ss t é t tr t é r tr st é t s 2 r s s q rr s t à s t t s s s s à sé t s t é s r r r s t t s q t é t tr r tr té r rs s t t s r s 3 rt s è s s è s t s r r tr t 1 s r 1 è s ér t t 1 r és r t s t s t t s t rt2 t s t t tts t t rr str t tt r t r t s
101 r t st ss q s è s rt s s è s t é é ç tr t rs s t rr r s r rt é t tr s s ts 1 r ss é q P r 2s r rt é t tr s 1 r ss t r é r r 2 t ès st à r r r t é r t 1 r ss s s tt 2 t ès tr té r s s ét s s s t é s r r è t t s r t s è s s s 1 s s t s t é ér t t stés r r é s ré s 1 r ss tr s ét s s s t r é s à t st r s séq s très s 2 t ès tr r t é r q 1 st r t rs tr 2 é t r 1 r ss t t t 2sé rré t tr r 1 r ss t t s r tr t 1 s 3 s r t s s t tré r t é r s t tr s 1 r s q st t è tr s 1 t s è t t s t é t tré q s è s é t s t à tér r s è s t 1 q tr t s ér s tr s è s q s t 3 s r t s 3 s r s P r ér r s s è s é t ç tr s t ré é t r 1 r ss à è s s sés tr s s s è s 1 r és str t r 1 r ss st s r 3 s s è s t s tr s è s q rt 2 t ès tr t t t st r q s s è s 1 r és é t s 1 ê s ç tr s rés t ts s r és s tt ét s t tr r és r ét s r q s tré r t é r tr r 1 r ss t t s r 3 s t s è s r s s r t rt s r s s s ér s t êtr t s st t st q s t sé s t2 s s t s t s t t t té à r t t t é ré s è s 1 s t t sé s é s 1 r ss é t 3 s r t s r t t t s r é r é t 1 ré t s è r t str t t é r r s t 1 s r 1 r ss s rés t ts s t t é rô rt é t tr 1 r ss s è s r t été tér ss t t st r rré t s r s é s t s r s rs q tr s ts r tr s s t s r à tér r s s t r à tér r s q st tr r r r r rré t t t q s t s s s t r é q très ré t t t s é s s r r tt s è
102 rt q s t r t q s t t str t ré tr s ér t s î s é r t t t t è 2 t ès r tt t t st r tr té é t tr s r t t t é rt à r ré t té t sé t q q s t s s tt ét r t t st r tr té é t 1 r ss r t s q t r rt è s é t ç tr t rt è s é t ç t t s é rt t s 2 t ès s é rt q s t s s r s t t t r 1 ré 1 st ré t tr s tr r tr s ét s t tré rt sé t st s t s r 1 r ss s s t t tr é q à s è s é t s t t q tt s s 2 t ès tr s ér t t à r é sé t st s t s t r ré 2sé s rs 1 é s 1 r ss ré é t és t s t s à é r è s tr s é t ré é t és tr ét s r tr s r t s rs s è s r s s q s è s s t s s sé t st s t s s sé t s t tr t sé t é t t 1 t r t r ér t è tr s t é t tré q s t rs tr s r t ét t s tr ts sé t st s t q rs s t tr r s st à r r r s rré t s 1 r ss rt s è s s t rs r t 1 s t t tré q 1 r ss s è s 1 és tr t s ét t rré és t ér t r r t q t t à s r ètr r t q s t r r è rt t s r s r rré t st q s r t rré t tr è s t st r q rt s t s t é s ré t ç t t tr rt ét t tr é r s t rs tr s r t s s t s tr t s é t s r r ré t q q s t s s t 1 r ss s è s t êtr s séq r ss r t s s q t s t ré t t t 2 t tt s è s é és r t t t ré é t é r ts s t sés s r s r s tr t 1 s très ré t s r t q st r tr t s r t t é é è r t t 1 r ss q s s q q q s s ts 2t q s r t t tr s r ts s
103 té r è s 1 r és st t s 2 t ès s s t r t t r s s s t è s r t rès s t rs r ré t r é t 1 r ss à t r t r t t è s s q r té r s tr s r ts è ss r à 1 s st ê q ss r à s t s à s è st ss t t st st q s s s é t q è q r s 1 r ss ç st st q r q r é t rt r r sé t s tr t s 2t q s s t é t r à st st té r è r t 1 q r s rs é è s ré é t s r és s ttér t r str t s tr s r ts s s r t q t q 1 r ss è é s r 1 r ss t q Y i = K K st t n i t n i r n ième è s 1 r é Y i ét t 1 r ss è i s è s 1 r é st K K s 1 è st 10 s r t é r tr r 1 r ss t t s r t t s t r t r 1 r ss rès rt t s r r t rt s s rés t ts s t r és 2 r st t s s s s s r rt sé t st s t t r t t rt é t tr q r 1 r ss s è s r r s s sé t s t 2 s t très tés 1 r ss r r té s è s é s s s r ss s t t r st t rs tr rsé st r q 1 r ss s è s s t rés t t éq r tr sé t st s t t ér r é r s tr t s 2t q s é r t r s r t P t êtr rr é s é s t é t r ttr t r s é t r ss t s é s tr s r t s t q s q t t r 2 r t r tt t s é t r t r tr s r ts q s ét 1 r ss 1 st t t s rs t s r ét r 1 r ss s è s P q t t t t s ré 1 s à s s ss t r s t q s séq ç t é t s t s r ét r 1 r ss s t s s r r ts P q t t t st à rt r s t s sq r t ét s t é q
104 r r str t è s st r t s s à t r sé s s s é s q r s s r èr s ét s s r r r è s s t é t s s à t r r r s s é s s r t s t q s séq ç t é t q r t s t t s s r 2s r t r s 1 t q s t t t r r r ss rt r s t s t é ts Pr s é ér 1 s s séq ç t é t s s à t s t 2 r t s é q é t r rq é r q t r r s r tt t q t t s è s s é s s t t é s r s s s é sé s s r s rt s s s r t té s à s tr s r ts tt s t à s é s tr s r t s séq és séq ç t é t r r s t r r t t s r ts t s à rt r tt t q t r ttr q t t s s r r s r s tr s r ts r trés st t é t ré ér q s t é séq é q 1 r ss q ré é t séq é s t s s s s t s rs r r s r r s tté r r t s s r s é s t t t s t s s st très tr sé t très ré q s t t 1 ér t t st t st q s r tt t très s q t t s s tr s r ts ré t é ér t ût s t s à r é t r t r q t t 2 r t s s t s s s à s rs s 1 r ss t êtr q t é q sq à rt r ss s q s t s t r s è s séq r r 1 s r s t s 2 r r s r s s t ss t s q t t 2 r t r sé ét st té 1 tr s r ts é à s t ré ér és s r tt t s ès à séq tr s r t q r t s ér r s r ts é q s ê r s s r s 2 r t t êtr té r r séq q t s t s é t tr r s t q s séq ç t é t s t r t s ê s s s t té s r q t té t t tr s r ts s t s q t tés r ét t t
105 r t tt t st é à q s t s s séq s r tt t st r s ér ts r ts st ss q s r t s séq s t t rt r rs s è s ré ér s s r é t t à èr s 2 r t s r sé s s r s s s r 1 t s rés t s s r r s r s tr s r ts tt s q r t é t r r s s è s r s s s s s s 2 r t r t tt t q rt èr t tér ss t é t s t q s séq ç t é t r tt t ès à séq s à rt séq tr s r t s tr 1 è ét s s t sé t q é t 1 r ss tt t q st t sé tr s r t q à s s é t r s ût q st é à s t s r ts ts s tr s r ts s é t st r rt r t r r séq r sé r tt t q q r t 2s s séq s t tt t q rés s r r r q t t s q s r ss t é ss té r s r r r s tr s r ts t q t é t s t sé tr s r t q s s q s st à séq r s r ts ss s rès s r ssés é t r t s s sé t ré s st t tt t q é ss t r s t s t s r t tr s r t r r q t t st r r r rt t s tés s séq rs t s st r t t q s q ès à séq s tr s r ts t à s 2s s ssé s r 1 è r è tt t r q st s té r sq t séq tr s r t s r s rt st r r tr 1 t é r t t r r r q t t s r t r rs s s s s t P r ût éq t r t été ré ér s r t r rs r ré t s é t s ré s r r 2s st t st q tt rr r t êtr sé r é r t r t st t st q r ét r ét rsq s s é ré tr 1 ér s t s t é t ét t q à rs t ts t 1 été r té r s r tér st q s s t s à t r tér st q s q t s t rt t é é t t t r r t r s t r r q t t
106 Pr 1 s ts t rts tt t ès t t q t tt t ès été très r ss t t s s té s s étr ts s st t st s rét sés r r t s r 1 t été é és s s r ts é és P r s q s ét t s t tt t ès s r 1 t s s s r r s t s t s s t q s séq ç r ré é 1trê t tér ss t à s t r t q t t 3 s é à ss 2é r r r t 1t s t s s s t t ts r rés t t tt t ès st t r s è s ét 1 t s é t s t s t s è s s s t é ér t sé t s t rt réé r s r ts és t q r s r â t sé t é à st t s é q rt èr st é r à rt r s é s rt t t é t 1 r ss t rt q é èr tr s é s 1 r ss t r 1 r ss s t ér t s q st st r é r q s s s ts 1 r ss r s è s r r és s r s r s s r t ts r s q s t û à st r s s è s à s é è s ré t
107 P rs t s r t q s q s r s r 1 s t t 1 ér t t 2 st r r s r r ts s s s q s r s é s ré sé s s s s 2 s sé r r s r è s ét s ét t r t és t q s é s t t q s q é ss r t q s t q s 2s tr s r t ét t r s à ér r t s s ét t très s à r s r t r s t s é s rt èr t r r s s é r t s s s t t s s 2s s r r s s s r 1 tôt q s q t s s s ts s t s é t s t t s r è s ré t és ût r P r rs s s s s tés q s str t t s à rt 2s 2 à 2s tr s r t q rt t 2 r 1 s ts t t 2s q s q r t st t r é t tér ss t é é r s t s s r t r t 1é q r r t été très tér ss t à q t r s ss à 1 r t r s ér s t s é t t é t s r s é t t st r rés st s é s r s t r é s 1 s t s s t sts rt té r s t s s s s r é s é s r ç s s t s s t r t ss s à t r q à t q r str t s 1 ér t t s ss s r t r t r é s ér t s t tt t ès r èr t rr t q s s rt s r rt té t s t t t r t r t r t s t t2 s rt à rt s st s t s t stér s r t r t s à t à q r st t t r t s t t s r t q r tt t rs é t trô é r r s s tr s t été t r é r s s ssé é t r t t s r r s
108 r s s t s ê é s é ts tér s t q s q t s séq s r s s r tt rt 1 ér st t s2stè t r ét t très tér ss t à s r r s ét t s t r r t t t r s s t t à t st r sé t st s r à q s s t s ré ss t t r t t tt ét q st rt r rés t t té s é t s s à s s t s t r s r q s r t t é ts t rs P s rs r t t s s t ss s t tér ss t s r rs r ét rés té s tt t ès tr s r t r tr s r t t rs tr s r t t r s r s 2s s tr s r t s rs t s s r é t 1 s s ét r rt t t t rt è s é t s s 2 t ès tr té q é ss t t r s t 1 s ér ts t s r r ér r s s è s 1 r és t t r ê ré s r s ts s t s 1 r ss s ê s t s r 2s s r s s P s rs ét s t tré rt s r s 1 r s s tr s s 1 s t t r t q s r t tér ss t à s r r s s t s ét r ç s é q rt s r s r s tr r s r é è rt r r 1 ét 1 t ét r s r s t st 2 sé t s 1 ét s r s r r t rs t rs s ét s t tré à q t 1 r ss ét t t ss s é q s t rt2 t tts t t rr tr t r st é t s t rr r s r rt r t r ét r s t r s t r
109 ét 1 t t t s t r s tr r t r ssè t t t q é s t r s s s ss s t r s r t tér ss t 1 r s ér ts è s è 2 s s t s r tr r st r é t s s s è t r r r è st r ê rr t ét r s ûts t t s s rt s s t s 1 q r s réq s s t s r s rr t é t s r é t s réq s é q s s rt s s ê s t s r s t 2s à q q s é s t r s r r s è t réq r q q s s t r t s t q 2 st rté r r t s t q é t s s st s r q r s ss t è s t rs s s s t à réq r s r t 2 t ès ût è s r t s tr t r s st s s rr s é t 1 r tr s è s ts ré é és r tr s r t q tr s 2t r s P q q s t t tr s ér s s r r t s st é é rés st t été é 1 r ss s tr s s r r ts t 2 s tt t r t s t s ét s st 2 t ès é s t s r t tér ss t s t r 1 r s rès 2 t ès s t é s t r q s s r sé r 1 q r s tr s r 1 r ss 1 r s rr t êtr t sé s r èr s st r t à é s r 1 r ss s t str t s s s s 2 t ès s t r t st t é t2 s ss 3 tr ts s r t s t s r t r s s s tt t ét r r s s r t t s séq t r 1 r ss s 2 s r t t st r tt 2 t ès s r tr s è t s t é s 1 r ss r t é t é s t 3 str s è q st t t ér é t à t q s s ér é s t ré t s s st 1trê t ré t q q s 3 s à t s é s q s r s s rr t s rêt r à s 3 t s r st rs s ss ss tr 2 t s q ré t t t q s é tt s st é s s
110 q s s t s 1 s è s st t t s 1 s s 2 tr s
111 r tt t t s rt P tr t s t ts s tt r r ss P s r s r t r P s t t r t t r s r s s ts s r 2 rs t s t s r t rs t r r t 1 r ss 2s s r s q t t rss 1 s t Pr t rs t2 Pr ss Pr t rs 2 tt P tr st tt r s 1 t s t r t r s st r r s s s r t tt t r t t r s t rts r r t s t r t2 s r r tst t r rr2 s P s t t t 2 t r t t 2 t 2 s rt t t rr s s tt r t t t r P st t tr s r t r s t st s t r2 r t t tr st 2 r s tt r s r s t s r t t r2 t s r s s s t s r t rt t 3 t 1 r ss 2 t r s t Pr t t q r r r t t t r s r 2 rs t s r s st r s s t s 2 t s r P P st P s r 2 t P t s 2s s 2 r s r r s s s P t r tr t s s r2 r t r t r r t t t st 2 t t s 2 s tt r tt t t r r s t st s 1 r ss t s r s r s r r r rr 3 r2 t r t t P r t r 3 s ts t ss t r t s
112 t rt Pr s r s t s t st r st r 1 t r s s rt s s t r s r ss s ss r t r t3 2 3 tt r P t r P 2 rs r r s st t r t r s st r Pr t ss t r t t s t t 2 2 s s s tr 2 t s r t s r t r r rt t t r 2 t ss t tr s r t r t 2 st r 2 r t t t2 t r s st r 2t r P 2 2 r t s t s s t r r s st t str s s t r P r s t t t st t st t s r r 3 t r t 1 r ss q 1 r ts r t s t s s t t s r r t s t t r s s s 1 st s s r 2 2 r st r t2 t s 2 P s r é t t 3 tr ss tr s s ts t r s r s t s r t2 Pt 2 P P t P t2 t r t2 r tr tr ts t r t s r s st r s s r r t s r s t ss s t 2 r s st r t r r P r st t t tt r r r s r 2 r s rt ts ss t t r s st r s st r tr t r 1 r ss sr t s r s rs s r r s s s 2 r s P r s P t rt s 1 r ss 2 r s t r 2 s r s r 2 r rr 2 2s s Pr t r P r 2 2 P P t t t t r t t r s r s t s 2 r t t t Pr t s t r r rr t t tr s r t r s s s t st r 2st r r ss str r t r t t s t r s s ts r r t r s r 1 t2 t 1 t t 3 2t r s
113 rr2 t r st t t r t s r s tr s r r t r r r r 2 t t r r t 1 r ss ss ss 2 st s t t r2 t r s st 2 r t t r t s r s st r r t s t str t r t r s s t r s st r ts s s s r s s s t s t3 rt r s r st t tt r P t r P s r t r2 ts t r r tr tr s s r s t t ss s 1 r ss t r s st r s t r s st 2 t s r s t r r r s rt 2 r t t s s t r s 2 2 t r st 2 t r r ss s t s tt r ts t t ss P r s r r 3 r s r st s 2 t t 1 r ss 2s s t t s s Pr t r s 33 s t t s t t r s 2 s t r s 1 s 2 t rr r s s t t ts s q s r ss t s 2 1 t t s r r s 2 r st r t2 t r s s ts r t s t s st t r P r t s s s r t t r s r rr 2 2s s tt r r t s r t s r s rr r r t st t s t tr t t r t r P t3 rs t P rr2 tt r P t s P ss t t s t r s st r s r r r r s 1 t s r s r t q t r t 1 t s r t s r t r s s s s st r s s P 2s s t 1 r ss r s 2 t t r t t t rt 3 t r ss t r r P t tr 2s s t r s r s s 2 s t r rr 2s Pr t
114 t r 2 t s t t t s t r s tr t t r r s s P 2 t r s é r t s r t ss t tr t tr s t r t r s P 3 s 2 t s t r s rs t2 Pr ss r t r rr 2 s r t 1 r ss r P 12 s s r t r2 r t s r s tr s t t 2s t t r t s P r t r2s s r t s r t s t s 2 t t st s 3 t r s s t s r r 1 s t 2 r2 t Pr t rs t2 Pr ss Pr t rs 2 r t t r 2 t r t r t s t r str ss r s s r s 2s s r t P rs t s 1 s s s 1 s 1 r ss t t r t P s ö r ss s P s t tr r r t tr t rs r t r t 1 r ss tt r s 2 t s t 2 s t t t tr s r s s s t r s st s t s 3 P tt rs s rt s t r r s r t s r t t 3 t s r t P s 2 s r 2 s r r s r t r t 2 tr t t q t t t t s rt t 2 t tt P t t r t r s t t t r t r t2 2 r s s t 32 s t 2 r s t s t r s Pt r s r s s t t r s r t t 1 r ss 2 st Pr t 3 t st t r 2 s q r rr 2s t r t s s s st r s P str s r rt r t t t t 1 2 s2st s t r r t t st r2 s t rs t2 t 2 s P r t s ss t2r 2 t 3 s tr s r t r2 ts tr t t t r r t tr s r t r s st r
115 s t r r r2 t t r t t t s 2 r r rs 3 t 2 1t s s 1 s t t2 1 r ss r s st r t s s t r 3 t t r2 t s st 3 t r2 ss 2s rt P rst t t P t s s r rr 2s r t t s q r 1 r ss r s s r t t 2 r s t 2t r P 1 r ss r s s ts P st P 2s t st tr s s t 1 t t t r 3á 3 t rs t P tr r t r t tr s s t t t r s st r P t r s s t s t r t r t r t t t t t t r st r 2 t 2 r t r t t t 2 r s t rst r r t 1 r ss s s r s st s s tr s r t tr r t t s t r 2 s r r str ss t s ts t r r t s t P 2s t q t r s r tr 1 t r rs t r s t s r s r 2r s t r r t t ss s tr s r s st r t s r t s r r r 1 2 t r t tr s r t r t t t s r t Pr t rt2 s t r t sq rt r r tt t t t st r 2 s 1 r t s r s t s rt2 t r t r s r tr t r t t t 3 s 2 r t t s t s s r s 1 r t s 1 st r t2 2 r s t t P t str t r r r t s r s s s r t s s r t 1t r t s rt 2 3 t ts t t t t s t tr t2 t s
116 s s t P rs r s t t r r s s r t s s P 2 t P t t s 2 t s s q r t t t t t r s st r s s 1 r t t s r r t s t t r s s s s t s ts s ts t s t s t 1 r ss r rr r t s P t s P r t s 1 r 2s s r t t s sts t ss s s s s t s 1 r ss r s t s r t 2st t t r t 2s s r sts s r t s r s r s t Pr t r t r t s r t t s t s t r t r s t 2s s r sts s s s 2r s 2 r á r s t P r t r t t tr s r t s r t t t2 ts t s t r r s t2 t s tt r s rr tt r P t r t t P rs 1 r ss r t r r t s r s st r 22t P 2 P t 32 t q t t t r t r r t s r s s s r t2 Pt r tt rstr tt r t q r t r t r t t t r s t t r s s s r r ts r t s t s s t r t r t 1 r ss t t r t s tr s r t s s t t s t 2 s r s r s s t r ss r t st s r2 t ss s t s s s 2 r t P P ss r r t s tr t s s 1 t2 t tr s r t r r s st r t t s t t t st r r r t s r t t r t r t s s rs r s t s r r r 2 3 t t s s s r t st s t r s st t t 2 r t2 s r t t2 t r s s s s r t2 Pt 2 t rs té r q t t s s r t 3 ï s 3 s r s s
117 s t s s r s s s r s st t st t t 1 t s s t s t t r s s t r 3 3 t r s st t r t r t r2 s q s r tr s s ts r s t rr s r tt s P rt 3 r t r t t st 2s s 1 r ss r rr 2 1 r t t r t t t st rr rt t r 2s s r r 1 r ss r rr 2 t t t P Pää t ss r tr t r2 1 r ss t s t P ss r t3 r r s r t Pää tr tr s r t t P r tr t r2 r t r rs t2 Pr ss r tt P 2 s r t rr2 rt r 2 t 2 t t s t r r t r s s s 1 s s t s r t st r r 2 r s s s s r s s q s P 2 t s 2 P r 2 r t r t s r r st r t 2 t t r s st r s r s s t 2 2 t t 2 r rs t2 Pr ss s r r s s t s r r st r r 2 t r s st r s s s r t t Pr t r2 2 P P s t t r tr s t s t s s t r s st r s s r r t 3 2 t s 2s s t r t s t 1 r ss r t 2 r s r s s s 2 r P r r t 33 t st t r rr 2 2s s 2t r P t s t r s st r s s t r 2 r r t P r st t t 2 r s t r s s r s st r s 1 P t s t r t2 t P r s t r s t r t r st r2 r s s s s t str t r t s
118 r tr s t r r s t t r t s sts r t s t str t r r s s s s t 2 s r t tr t st s r s st r s á r s t rt t s r 1 r s s r s r t s s st 3 s t r t2 t t r s t tr t s t Pr t2 t t st s ss 2s r r t t r rs t2 Pr ss r r P t s è t s 2 s P r s rt r r t t s t ttr t ss r s t t r s P Pr s r t r 1 r ss r s r st r s r t r s ts t ts r r r rt P r r tt r P t r t t t tr s r s s t s r s t s P t r tt s t r t 1 r ss r st t s t r t tr t s tr s r t tr s t r t t t 2 r s r t t r r ss t s r ss s tts t2r t r2 s t t st r s t t r t r2 2 P P s r r t r s t P 1 s 1 r ss s 1 t r t s2st r s t s t s q ss ss t t r r t2 r s t 1 r ss rr 2s s s 2 P t Pr s r s r s t ss t t t r s s t r s P ts 2 s t 2 r t r t s P P t t st r t st t r r r s s P 2r 2st t s t r s s rs t2 Pr ss r rt t r st t r s st s rr s st rr rt t r st t ss t t s t r s st ss t 2t r P 2 P st r tt t st t s t t s t s s s t r s s s 1 r t s s q t P
119 P rs 3 t rt t s 1 r ss r s Pr t t s r t t 1 r ss st rs t r s tr s r t P s t 1 r ss 2 r s r s t s P t r tt r s 1 r ss r s r s s 2 r s P t r ss t s 1 r ss t st r t2 2 r s r s s s r t r 3 P t r tt s r r t s 2 s r t r2 s t 1 r ss r t t t st s s s r t r t2 r s t 2 t s s t 2 P s 2 r t r t r tt r s 1 r ss r s r s s 2 r s 1 t t s s 2 r 3 t s t r s 2 st s r 2 2 r s t t 2s s r t 1 r ss t r r 3 2 t t r s r t t r s r s s t t2 r s st r t s r tt r t 2s P t r t t P rs P t tr s r t s r s st r s s st rs t 2 t r s st r s 1 st r st r t t r s st r r s s s r r s r s 3 s t 3 r sst t r s s s t t t str ss s r s s 2s s 1 r ss 2t r P s r 2 2 r rr 2 P t 2 r s r s r s 1 r t t s ts s 1 r r 3 2 r t t2r tt r t 1 r ss r t s q r s s r t t s tr s r t s 1 t t t r2 s r t t r t s 1 r ss t t r t r t t r s t P s r t r r r t 1 r ss t t r t s t t
120 s t r r t r r t r t s 1 r ss s t r t s t t rr t r 1 st s s t t s s 2 r st r t2 t r s s s r t 2 r 3 t t s rr t r r t st rt 2 r s t t t s s s r s s s r t s s t 2t t s r rr 2s t st 2 1 r ss s 2 r t s s r t s P r t ts r t r2 r ss s r t t r2 r ss s t r2 r rt r r P tt 2 t 3 t r t t ts rts str t s tr t s r s t ss r s t P 2s P t t t t r t s tt r r ss r t2 P r t2r r t P tt r tr s r t r r t r2 s t r 2 tr r t r s st t r s Pr t P r r r t 2r r t tr s s t s rt s 1 r ss t r ts P P r 3 s t t s r r t s t t r s s s r r ss st r t r 3 t 2 r st r t2 2ss s s t s Pr s r s 1 r s s s t r s r s t Pr t r s r t s P Pr t r r r s 1 r ss Pr t Pr t rr r r s r t r2 t Pr t Pr s ss r s r t rr t r t t r s r t r2 s tr t r r t 2 t t ss r t ss tr t r t t s st r s s t s t r t t P r r t P s Pr t r t t2 s 2 2 t s ss t s t t t t t 2s s 3 r r tt ss r t s r r t r Pr s t t r s st r s s r P 2 r t P st r r t t r r st r s s t sq t 1 s P s r s
121 r t t s r q r r r s r r r t s s 3 tr r s t 3 t s r s t st s t 2t r P s r 2 r t t t t s 3 t t t r r t s t s rr t r t r t r2 2 s t t r r r t s t t t s r s t st r rt r s 3 r s t t t P t 1 r ss t r s st r s r t t 2 P t st t s 3 t t r s st r s s 1 2s r t 2s s Pr t s t s s r 3 t t r r t 1 r ss 2s s s q t r tt t r 2 r t r2 r t r 2 s r s P t tts t t rr 1t s t t2 1 r ss r t t s s s s t s s t s 3 t t t st s q t t t st t tr s t s ss t t tr s s s rt r s s s Pr t t t rr r 2 t t3 rt r P r tt r P t 2 r r t tr s s ts t r r t t t t t 2 s P t ö t ött r r P tt r s r s t t r t2 r s s t s s t r q 2 t t s r s s ö t ött r r r s t t r t r t t r r t s r s s s r s r r t r r s r t 2s s r s s ts r rst t rs ts s r s st r s r 2 t tr t r t t t t tr s r s s t s r ss t rs t 32 s r 2 Pt r r2 t t r s t r2 t s r s t r 2 r rs t2 Pr ss r
122 t t 1 t s t r s t 2st t s st t r r t Pr r st t t r s st st s s t ss s t r t r2 t 2 t r r t t t r 2 2 tr t r t 2 1 r ss r t t t s t t rt t s s s 2 r s r s st r Ps2 t t r r r r 2 t r s s ts s s s s 2 s r t r2 t s 2 Pr t rt t r 1 r t s r t st rt ss t t r r t s t r s Pr t r t t r t s t r s st r r s t t 2 r s 2t r P s r 2 r s st r t t tr 1 r 3 t 2 2 r Pr 2 P t r r t t r r t t 1 t2 r s st r s t r t r 2 r t t r t s 1 s t rts s r s s ts s s s t 2st s P r t rt P t t r t 1 r ss s r r s 2 P t P r2 r s t r r r t 2 s r s t 2 t t s s s t s ss rst 2 2 P tt t t P s r t s t t s r t 1 r ss t t r s t 2 2 r2 r t r t st r t2 t t t s r s st r s s t s t r s rt t t s 3 t st r t 1t t t s ss ts s P t s s 2 r s t s t s 1 r ss s t r s st t s s t s str s st t t 2 r t r s t 1 s r s t
123 str t3 r r t r P r t rr tr st t 1 r ss r s t t st s t s s s s s t s s s t ä t r s s r t r t2 r r rs P r r t 2r r r s s ts s rt tt r t 1 r ss t s t rs t P ss t t t r r t tr s s t r t 1 r ss s t r s st ss t P r s Pr t t t r r r t t ss s 1 r ss t r t s t t r r tr t r t 1 r ss Pr t t t r r r t t s s 1 r ss r t r r t t r t t r t tr s r t s t s s s t t r s t t 2 P r 3 r t r P t2 r t 1 r ss tt r s Pr t r s t t r t r s t s 1 r r s st r s t P 2s s r t r t r s s t r r r t s s st 2 t t r t r t t t t2 s t tt P r t r t r2 s s tr s r t t r tt P r t r t ts s tr s r t r2 r t 1 r ss r s st r t s tt P r t r t r2 s r 2 1 r ss r s t t r s s s t t 2 r s st P t t3 t r s 1 r ss r t t r t tr s r t s q t r s t s r t rr r ts 2 r 3 t t r r P rtr t r t r tr rs r t ss r2 1 r ss r s s t P 2s t t tr st s s t q 1 r ss tt r s r s r t s s
124 r 2 t r2 s s r t r2 t t s t t t s s t t t r t P rr t t s 1 s r s r 2 r rs r s r r r2 r r 2 t t r P t tr s r t s s ts r r s s s r s s t r 2 r s t r P q tt 2 r s t r t r rr 2 s 2s s 1 r ss r2 r s t 2t r P s r 2 r s s t t r P r r r t s s ss s 1 r ss r t s 1 2 r s s ss ss t r t 2 P t s st r r st r s t r s r tr t t st r r r st s t t r P r s r t r str t t r r s 1 s 1 r ss t s r s t r
125
126
127 1 s s s 1 s s tr r 3 r èr t s t t s r q st rs ré r t t st êtr t s tr r 3 é t st s tr t s r s t é r t s rs rès s q s é s st rs rés tés t t ès s tr r 3 s st s è s ér t t 1 r és tr t s Prés t t s r s s é r r s r tt s t r s r tt t r t s t r s P st rs t r t r 2 ût P r s s s t r s r s t r r t2 t r2 2 r ût r t2 r r 2 t r t 2 r t2 t r2 2 r ût ü
128 Genetica DOI /s Population transcriptomics: insights from Drosophila simulans, Drosophila sechellia and their hybrids François Wurmser David Ogereau Tristan Mary-Huard Béatrice Loriod Dominique Joly Catherine Montchamp-Moreau Received: 22 November 2010 / Accepted: 7 March 2011 Ó Springer Science+Business Media B.V Abstract Sequence differentiation has been widely studied between populations and species, whereas interest in expression divergence is relatively recent. Using microarrays, we compared four geographically distinct populations of Drosophila simulans and a population of Drosophila sechellia, and interspecific hybrids. We observed few differences between populations, suggesting a slight population structure in D. simulans. This structure was observed in direct population comparisons, as well as in interspecific comparisons (hybrids vs. parents, D. sechellia vs. D. simulans). Expression variance is higher in the French and Zimbabwean populations than in the populations from the ancestral range of D. simulans (Kenya and Seychelles). This suggests a large scale phenomenon of decanalization following the invasion of a new environment. Comparing D. simulans and D. sechellia, we revealed 304 consistently differentially expressed genes, with striking overrepresentation of genes of the cytochrome P450 family, which could be related to their role in detoxification as well as in hormone Electronic supplementary material The online version of this article (doi: /s ) contains supplementary material, which is available to authorized users. F. Wurmser (&) D. Ogereau D. Joly C. Montchamp-Moreau Laboratoire Evolution, Génomes et Spéciation, CNRS UPR9034 Avenue de la Terrasse, Gif-sur-Yvette F Cedex, and Univ Paris-Sud, Orsay, France francois.wurmser@legs.cnrs-gif.fr T. Mary-Huard Statistics and Genomes Team, INRA UMR518/AgroParisTech, 16 rue Claude Bernard, Paris, France B. Loriod U928 INSERM, Parc scientifique de Luminy, 163 Avenue de Luminy, case 928, Marseille Cedex 09, France regulation. We also revealed differences in genes involved in Juvenile hormone and Dopamine differentiation. We finally observed very few differentially expressed genes between hybrids and parental populations, with an overrepresentation of X-linked genes. Keywords Expression differentiation Population structure Ecology CYP450 Hormone regulation Drosophila Introduction Genetic variation at the sequence level has been widely studied within and between species, to identify the forces that drive evolution (e.g., natural selection, genetic drift). The interest has been now turning to changes in gene regulation leading to genetic isolation, and ultimately speciation. Indeed, gene expression can have a strong influence on downstream phenotypes, and therefore its variation is likely to be a target of natural selection (Pavey et al. 2010). Large-scale technologies such as microarrays provide genome wide information that can be used to assess expression evolution (Gilad and Borevitz 2006). Several comparisons are possible: among populations of the same species, among species, as well as between interspecific hybrids and their parents. Natural populations have been previously studied for regulatory variations in several organisms from yeast (Townsend et al. 2003) to hominids (Storey et al. 2007). Two studies have reported differences in gene expression between African and European populations of Drosophila melanogaster with respect to sex biased genes (Meiklejohn et al. 2003) and genes involved in toxicity resistance or flight musculature which are potentially involved in adaptation (Hutter et al. 123
129 Genetica 2008). No evolutionary studies so far have compared expression between closely related species and their hybrids using a set of distinct parental populations (contrasting with studies based on a single strain for each population or species). This is the approach we have adopted in the present work, studying closely related Drosophila species belonging to the melanogaster subgroup. It will allow us to assess at the expression level, not only the extent of population structure but also the importance of misregulation and additivity in hybrids. We used here different populations of Drosophila simulans and a population of the sibling species Drosophila sechellia. Drosophila simulans and D. sechellia have long been thought to have separated about years ago (Hey and Kliman 1993; Kliman et al. 2000; Lachaise and Silvain 2004), but more recent data pointed to a split occurring around years ago (McDermott and Kliman 2008). On average, the DNA sequence divergence between these two sister species is around 1.5% (McDermott and Kliman 2008). While these two species are phylogenetically very close, they are ecologically strongly different. D. simulans, which originates from eastern Africa or Madagascar (Lachaise et al. 1988; Lachaise and Silvain 2004; Dean and Ballard 2004; Kopp et al. 2006), is now a cosmopolitan generalist. D. sechellia is an endemic specialist in the Seychelles islands, and breeds exclusively on Morinda citrifolia, a plant highly toxic for other drosophilids (including D. simulans) (R Kha et al. 1991). According to Oleksiak et al. (2002), gene expression differences between populations arise mainly from genetic drift (they did not show more differences within population than between). Our design already takes into account the variation within populations, thus differences shown there may equally represent drift or adaptation. Notably we expect a higher expression variation in D. simulans because of the higher diversity of environments occupied by the species compared to D. sechellia. The geographic structure of D. simulans has been studied using nuclear gene sequences or microsatellites (Hamblin and Veuille 1999; Schöfl and Schlötterer 2006; Baudry et al. 2006). These studies have revealed a differentiation between east African populations, other African populations (notably Zimbabwean), and European populations. In contrast, D. sechellia harbours little nucleotide sequence variation, which ranks this species as the least genetically diverse drosophilid (Legrand et al. 2009). Previous studies which have examined divergence in gene expression between D. simulans and closely related species, have used samples isolated from a single isofemale line (i.e. consisting of the offspring of a single wild-caught female) (Michalak and Noor 2003; Haerty and Singh 2006; Moehring et al. 2007). Thus, they have not taken into account the intraspecific variation. An exception from this is the recent parallel studies of sequence polymorphism and expression of six D. simulans lines originating from five different locations within the species range (Holloway et al. 2007; Lawniczak et al. 2008). The authors revealed interesting insights into the evolution of regulatory sequences between populations. However, each of their population was represented by only one (or two in the case of Madagascar) line, thus they could not take into account intra-population variation. The present study includes four populations of D. simulans each represented by four isofemale lines, four isofemale lines of D. sechellia and four hybrid populations (crosses between D. sechellia males and D. simulans females). This design allowed us to take into account two different types of variances: intraspecific variance (between the different populations of D. simulans), and intra-population variance (between the different lines which represent biological replicates within each population). We considered the four isofemale lines of D. sechellia as a single population as this species does not show geographic structure (Legrand et al. 2009). We used males because they are the most affected sex in Drosophila hybrids (since they are heterogametic they show more hybrid breakdown), therefore suitable to highlight differences linked with species divergence. Regulation breakdown in hybrids can occur due to incompatibilities between alleles at a given locus It can also result from negative epistasis between loci according to Dobzhansky-Muller s model of hybrid incompatibility (Dobzhansky 1936; Muller 1942). Differences in hybrid expression can also simply result from regulation divergence between genomes. Hybrids have been previously shown to harbour strong regulation breakdown compared to parents in different organisms (Gibson et al. 2004; Haerty and Singh 2006; Moehring et al. 2007). However, other studies have shown large scale additivity in the patterns of expression of the hybrids (Hughes et al. 2006; Rottscheidt and Harr 2007). These contrasting results may be caused by methodological differences (organ-specific vs. whole body/ differences in microarray platform) or by different degree of divergences and inbreeding between species (Rottscheidt and Harr 2007). These two non-mutually exclusive hypotheses will be considered here. We extracted RNA from whole body males to consider only general differences, and not tissue specific differences. Our populations of D. simulans were from Zimbabwe, Kenya, the Seychelles (the last two likely represent the ancestral range) (Lachaise et al. 1988; Lachaise and Silvain 2004; Dean and Ballard 2004; Kopp et al. 2006), and from France (a derived population). This approach allowed us to determine to what extent gene expression shows geographic structure in D. simulans, as well as differentiate between population effects (possibly linked with recent invasion) and species divergence. We observed a population structure in D. simulans, and consistent expression divergence 123
130 Genetica between D. simulans and D. sechellia. We notably observed a strong involvement of the Cytochrome P450 gene family, as well as genes regulating the juvenile hormone (JH). Materials and methods Drosophila stocks We studied four populations of D. simulans. Each population was represented by four isofemale lines (biological replicates). The populations came from France (the Rhone Valley, collected in 2003), Kenya (Nairobi, collected in 2001), Zimbabwe (Mazoe, collected in 1997) and the Seychelles Islands (Mahé and Praslin, collected in 2003). The four D. sechellia lines also originated from Mahé and Praslin in the Seychelles archipelago (collected in 2003). RNA samples The lines were mass reared in uncrowded culture, on axenic standard medium at 25 C, with a natural light cycle. For each isofemale line, C6 replicate cultures were raised in vials containing each 8 males and 8 virgin females (10 of each for D. sechellia). For a given D. simulans population, four different crosses were performed, each between females of one of the line and males from a different line of D. sechellia (at least 3 replicates per cross). We thus obtained four populations of hybrids corresponding to the four D. simulans populations (Table 1). The experimental design thus included a total of nine populations, each consisting of four biological replicates. Virgin male offspring from at least 3 replicate vials were collected within a few hours of emergence to create pools of 25 individuals that were transferred into fresh vials. Seven days later, each pool was frozen at -80 C. We used the Nucleospin RNA II kit from Macherey Nagel to extract the RNA from the pools of 25 whole-body adult males, yielding to a total of about 3 lg RNA to hybridize on the arrays. RNA was then reverse transcribed in presence of alpha d CTP p33. Arrays The arrays were nylon filters spotted with long amplicons from the species D. melanogaster. They were hybridised in the TAGC platform in Marseille. There were 7,041 spots: 5,931 different whole cdna of D. melanogaster and 1,100 control spots, either negative, or positive controls (a cdna of Arabidopsis thaliana: chlorophylle synthetase). cdnas were cloned into a vector, amplified and spotted on the array. Each spotted fragment contained both the cdna and a specific part of the vector for spotting normalization (first hybridisation). These arrays were hybridised twice. Firstly, hybridisation was performed with a P33 labeled oligonucleotide probe specific to the vector sequence spotted. As every molecule spotted contained this sequence, the radioactive signal (vector hybridisation signal) read was proportional to the quantity of spotted cdna. Secondly, after deshybridisation of the vector probe, we proceeded with the hybridisation of the cdna samples. A second radioactive signal was read. It will be further designated as complex hybridisation signal. Data normalization The normalization procedure was defined by the manufacturer of the arrays. Every spot for which the vector signal was smaller than 5 times the negative spots median was eliminated from the analysis. For both signals, background (measured by the median of negative spots) was subtracted. Inter-spots/intra-arrays normalization was then performed by dividing the complex hybridisation signal by the vector hybridisation signal. The last normalization step was to divide the obtained expression value in each array by the corresponding median of all spots (or by the median of positive controls), effectively normalizing the signal between arrays. The two approaches (median of controls/ median of all spots) led to the same results. Normalization quality was assessed visually by MA plot and box-plot of normalized expression values for each array (Supplementary Fig. 1). All genes with four or more missing data throughout the 36 arrays were discarded. Statistical analysis The statistical analysis was based on the whole set of 36 arrays. To determine differentially expressed genes, an Analysis of Variance (ANOVA) model was fitted for each gene (Kerr et al. 2000, 2002). The fitted model was the following: Y ij ¼ l i þ E ij ; where i is the population index (i = 1,,9: 4 D. simulans, 1 D. sechellia and 4 hybrid populations ), j is the biological replicate index (j = 1,,4), Y ij is the normalized signal (log-transformed), l i is the mean expression for the gene in population i and E ij is the residual variability. This model assumes a common variance for all populations, that is consistently estimated with 36-9 = 27 degrees of freedom for most genes (a few genes have missing values due to normalization). The homogeneity of variance between groups was verified using Levene s test (Levene 1960). We showed a variance homogeneity for all genes but 20 (FDR = 0.1, see Supplementary Fig. 2 for the distribution of P-value). We tested the equality of mean expression between D. sechellia and D. simulans 123
131 Genetica Table 1 Experimental design to obtain hybrid males 16 hybrids were obtained from crosses of D. sechellia males with D. simulans females. Each isofemale line of D. sechellia was involved in a cross with a different isofemale line of each population of D. simulans populations as well as between D. simulans populations. As for the comparisons between a population of hybrids and its two parental populations, we performed tests to compare the mean expression of the hybrid to the mean expression of each parental population, and we also compared the mean expression of the hybrid to the average of the mean expressions of the two parental populations. All these comparisons were performed using usual contrast t tests within the ANOVA model. For each comparison, raw P-values were adjusted by the Benjamini-Hochberg method, which controls the false discovery rate (FDR) (Benjamini and Hochberg 1995). We used a FDR of 0.1. A mixed-effects model does not suit our analysis, since estimates of some parental effects would only have been from two values. However, we assessed the effect of our analysis assuming correlated data by simulating data with parental effect. We did not find any increased number of false positive even with a high biological/technical variability ratio, the only consequence was a decreased power (Supplementary Fig. 3). Patterns of inheritance: additivity, dominance, overdominance To assess patterns of inheritance, we analyzed the distribution of dominance effects using the ratio d/a, where a is half the difference in expression between the parental populations (D. sechellia and respectively each population of D. simulans), and d is the expression difference between F1 hybrid and the parental average. If d/a = 0, it means perfect additivity (d = 0), if d/a =1, complete dominance and if d/a [1, overdominance (Falconer and Mackay 1996). We performed this analysis for our four parental populations, only to those genes which were differentially expressed between the parents, to avoid any bias due to equality of expression between the parents. Variance comparison To compare the genomic variability in two populations A and B, we propose the following test. This test looks for an excess of genes with higher (or lower) variance in one population relative to another. For a given gene g, we note r 2 g;a and r2 g;b the gene expression variances in populations A and B. If population A harbours more genetic variability than population B, then, for most of the genes, the ratio R g ¼ r2 g;a will be higher than 1, whereas R r 2 g should be higher g;b than 1 for roughly 50% of the genes if the two populations are comparable. Therefore a test can be based on the number N AB 1 of genes for which the empirical ratio R g of gene g is higher than 1. We note p AB the true proportion of genes for which r 2 g;a [ r2 g;b. We test H 0 ¼ fp AB ¼ 1=2g(A and B are comparable) versus H 1 ¼ fp AB [ 1=2g (variability is higher in A). N1 AB has a binomial distribution BðG; p AB Þ with G the number of genes. The P-value of the test is thus: PðN AB 1 [ n AB 1;obs jp AB ¼ 1=2Þ: This analysis takes into account the number of lines available to estimate each variance. We simulated data with different number of lines (n = 2, 5, 10 and 50) with equal population variance, to measure the impact of a small number of lines on variance estimates. This did not affect the P-value distribution and thus the error. Significant tests were around 5% (Supplementary Fig. 4). Further simulation showed this only affects the power of the test. Gene ontology Lists of differentially expressed genes were examined for statistical over/under representation of Gene Ontology 123
132 Genetica (Ashburner et al. 2000) terms using FuncAssociate (Berriz et al. 2003) with a reference background consisting of all genes in the arrays. Our array itself was compared to the whole genome, revealing several ontology biases in the construction of the array. This made essential the use of our array as background when examining differentially expressed genes for gene ontology bias. To further explore the terms and the corresponding genes, we used the Gene Ontology database provided by the Gene Ontology consortium (in May 2008). Results The experimental design allowed us to perform multiple comparisons, within D. simulans, and between D. simulans and D. sechellia, as well as their hybrids. By maximizing the biological source of variation (using biological and not merely technical replicates, Altman 2005) in populations (and species when applied), we revealed strongly significant variations. After all gene filtering during the normalization process, we assessed expression for 4,398 genes, which is about a fourth of the Drosophila genome. The differences observed are thus non exhaustive, but their consistency between all populations, through our large sampling, is supported by the power of the cross-design. Comparison between populations of D. simulans We did not detect any significant difference in gene expression between the three African populations. Contrasting with this result, all the comparisons between the French population and each of the three African populations showed differential expression. Respectively 6, 7 and 13 genes were found to be differentially expressed between the French population and the Kenyan, the Seychelles and the Zimbabwean populations (Supplementary Table 1). Six genes were differentially expressed in two pairwise comparisons. No gene was differentially expressed in all three pairwise comparisons. Out of the twelve genes which were over-expressed in the French population compared to at least one of the African population, four are cytochrome P450 genes (significantly over-represented, Fisher s exact test, P \ 0.05). We compared variability using a binomial test based on the fact that variance ratios of genes are expected to be half of the time above 1 under the assumption of similar variances, using only pairwise comparisons. P-values of the binomial test are provided in Table 2. The variance from the French population is significantly higher than the variance from any other population but the Zimbabwean (P \ 0.005, Bonferroni corrected threshold). In terms of variance, we observe a differentiation of the French and the Zimbabwean populations compared to other African populations (from the zone of origin of D. simulans). All other pairwise comparisons revealed significant differences, even though there is a wide range of P-values. It is important to note that this analysis is independent from the test of variance homogeneity gene by gene performed with Levene s test. It is possible to have homogeneity gene by gene, whereas on a global scale, heterogeneity can be observed. Drosophila simulans vs. Drosophila sechellia The comparisons between D. sechellia and D. simulans yielded to 347, 337, 353 and 518 genes differentially expressed with the populations of Zimbabwe, Kenya, Seychelles and France, respectively. Details of over/underexpressed genes are shown in Table 3. The striking result is that 304 genes are consistently differentially expressed between all four populations of D. simulans and D. sechellia (Supplementary Table 2). We can therefore assume that these genes present constitutive expression differences between the two species. Table 2 Above the diagonal: P-values of binomial tests under the assumption of equality of variance between the populations Below the diagonal: direction of the variance change F France, Z Zimbabwe, SimS Seychelles, K Kenya, Sech: D. sechellia * significant (P \ 0.005, Bonferroni corrected threshold) 123
133 Genetica Table 3 Number of genes over-/under-expressed in D. simulans compared with D. sechellia Population Total Over-expressed a Under-expressed b Zimbabwe Kenya Seychelles France a Genes over-expressed in D. simulans compared with D. sechellia b Genes under-expressed in D. simulans compared with D. sechellia Five terms were consistently over-represented in the subset of genes under-expressed in D. sechellia compared to every population of D. simulans. The molecular function electron carrier activity and the cellular components vesicular fraction and microsome refer to cytochrome P450 genes, as was assessed by examining the intersection of the genes with this annotation in D. melanogaster, and our set of differentially expressed genes. The two other terms (namely lipid metabolism and hormone catabolism ) refer to three genes: Juvenile hormone epoxide hydrolase 1 (Jheh1), Juvenile hormone epoxide hydrolase 3 (Jheh3) and Dopamine N-acetyltransferase (Dat). These three genes are highly pleiotropic as they are directly involved in the regulation of key hormones: Juvenile hormone and Dopamine (DA). Jheh1 and Jheh3 are involved in JH regulation by degrading it. According to Gruntenko and Rauschenbach (2008), the JH titre can be assessed by the JH degradation level. Thus, we can assume that an over-expression of Jheh1 and Jheh3 in D. simulans compared with D. sechellia implies a lower JH titre in D. simulans. The gene Dat is involved in the degradation of DA. Hybrids vs. parental populations Expression in male hybrids was compared with males of both parental species, and the mean expression of the parents. By this process, the differentially expressed genes were those different from the parents and therefore not showing the dominance of a parental allele on the other. This also excluded genetic additive effects since their expression had to be significantly different from the parents mean. A significant difference could be due to underdominance, i.e. failed interaction between the two alleles, or misregulation through negative epistasis. We found few genes perturbed in hybrids. No gene disruption was detected in hybrids obtained with the populations of D. simulans from Kenya and from Seychelles. Significant over-expression was detected for four and five genes in hybrids offspring of the French and Zimbabwean populations, respectively (Table 4). X-linked genes were significantly over-represented in this set of genes (Fisher exact Table 4 Genes over-expressed in hybrids compared to parents Offspring of F D. simulans 9 D. Sechellia Cp110 CG14785 CG4558 Es2 Z D. simulans 9 D. Sechellia Cp110 sm r-cup CG3795 CG31108 test, P \ 0.001). One gene was common in both comparisons (Cp110). This gene is involved in centriole replication, although its precise function is unknown (Dobbelaere et al. 2008). Another gene (r-cup) is involved in male meiosis, and its disturbance could have a role in hybrid sterility (Barreau et al. 2008). We also examined patterns of dominance and additivity. For this analysis, we included only genes for which expression is different in the two parental populations. We used the ratio dominance over additivity (d/a). About 45% of genes showed additivity to partial dominance (0 \ d/ a \ 0.5), about 26% showed partial to complete dominance (0.5 \ d/a \ 1), and about 29% showed overdominance (Fig. 1). Discussion Our multiple comparisons of transcriptomes revealed three main features: 1-geographic differentiation in D. simulans; 2- expression divergence between D. simulans and D. sechellia (about 7% of the genes), notably cytochrome P450 genes, and genes involved in hormone metabolism (JH and DA); 3- only eight genes misregulated in hybrids among which X-linked genes were over-represented. Assessment of the use of interspecific array Gene localisation No genes were found differentially expressed with offspring of the Kenyan and Seychelles population F French, Z Zimbabwean We used arrays carrying cdna from D. melanogaster. The use of interspecific arrays has been shown to introduce a bias in comparisons, due to differential hybridisation caused by sequence divergence (Gilad et al. 2005; Oshlack et al. 2007). However, D. melanogaster, one of the closest sister species of D. simulans and D. sechellia, is approximately as X X X X X 2R X X 3R 123
134 Genetica Number of transcripts France Zimbabwe Number of transcripts Seychelles Kenya d/a d/a Fig. 1 Distribution of the ratio d/a for hybrids crossed from our four different populations of D. simulans. When d/a = 0, d = 0, we are thus showing perfect additivity. From 0 to 1 (or -1), we shift from complete additivity to complete dominance. Over 1 (under -1), we show overdominance divergent from D. simulans and D. sechellia. The divergence between D. sechellia and D. simulans is thought to have occurred around years ago, while the divergence between the two species and D. melanogaster dates back 2 to 3 million years (Lachaise et al. 1988; Hey and Kliman 1993; McDermott and Kliman 2008). We can thus assume the hybridisation signal to be similarly affected by the use of a D. melanogaster array. Moreover, in order to limit the possible bias, we chose a long cdna array rather than oligonucleotide, which limited the effect of possible mismatches in the sequence. To assess whether there was a hybridisation bias, we looked for a correlation between: 1- the sequence divergence difference between each species and D. melanogaster and 2- the mean differences of expression between the two species. We assessed this in a large subset of genes included in the array (2,303 randomly chosen genes), using the sequences from the database ( We observed no correlation (Spearman correlation coefficient: Rho = ; P = ). D. simulans and D. sechellia diverge by less than 2%, which is comparable with the level of divergence of disparate populations of D. melanogaster. 80% of divergent bases with D. melanogaster are shared between D. simulans and D. sechellia (Dworkin and Jones 2009). In this study, they found little evidence of any bias between D. sechellia and D. simulans. Although on a global scale, the influence of mismatch is likely negligible, we agree that individual genes can be affected. However, Mezey et al. (2008) showed that the effect of sequence divergence is mainly an increased variance leading to a decreased power of the test. Our tests are thus likely conservative. To assess a possible bias, we used sequences from Flybase to calculate the difference in coding sequence divergence (D. melanogaster vs. D. simulans minus D. melanogaster vs. D. sechellia). This was calculated first for differentially expressed genes between D. sechellia and D. simulans, and second for the random set of genes previously mentioned. Differentially expressed genes are represented by a subset of 232 genes taken from our list of 304 genes consistently differentially expressed between D. sechellia and D. simulans. We could not consider all genes because of incomplete/erroneous ortholog annotation in Flybase. However, in both sets (random and differentially expressed), genes were sorted the same way. Although the P-value is relatively low, the mean difference in divergence between random genes and differentially expressed genes is not significant (Mann Whitney, W = 286,527, P-value = ). We assessed further the extent of the possible bias using a Q-Q plot of the distributions (Supplementary Fig. 5). This revealed that about 30 genes out of the 232 are biased toward a higher divergence compared to the random genes distribution. This bias can have two explanations. The first one is that 123
135 Genetica sequence divergence affected hybridisation, leading to false positive. The second explanation is biological: genes with a higher coding sequence divergence can also be genes evolving faster in terms of expression changes. Although there might be some bias, cytochrome genes as well as Jheh1, Jheh3, and Dat show a divergence difference below 1.3%, situated in the unbiased part of the Q-Q plot. This bias may have two consequences: a decreased power for the test, and a slight increase in the false positive rate. Differentiation between D. simulans populations Expression differentiation D. simulans originates from Eastern Africa or Madagascar (Lachaise et al. 1988; Lachaise and Silvain 2004; Dean and Ballard 2004; Kopp et al. 2006). This ancestral area is consistent with several observations of our study. We would like to point out the difference between the African populations of D. simulans, and the French population which are revealed with all comparisons. Each of the African population revealed around 350 expression differences with D. sechellia, while the French one revealed 518, thus showing a significantly stronger differentiation compared with D. sechellia. The direct comparison revealed no differential expression between African populations of D. simulans, while we detected several differentially expressed genes between the French population and each African population. This is consistent with the weak but existing population structure observed on microsatellites by Schöfl and Schlötterer (2006) and on nuclear genes by Baudry et al. (2006). Gene flow should be higher between the three African populations than with the French population, as expected from an isolation by distance model and the evolutionary history of the species. A low level of existing geographic differentiation was also described for morphological traits (Gibert et al. 2004). Among African populations, the study of Schöfl and Schlötterer (2006) shows a differentiation between sub- Saharan populations and non sub-saharan populations. Within the sub-saharan population a differentiation was observed between the lines from Zimbabwe and Malawi on the one hand, and the lines from Uganda on the other hand, the latter being geographically closer to the likely region of origin of D. simulans. Our study suggests a similar differentiation between the Zimbabwean populations and the population from the ancestral zone. Indeed, when we compare F1 interspecific hybrids with their parents, the Zimbabwean population leads to 5 differentially expressed genes (Table 4). This result is comparable with what is observed for the French population (4 genes differentially expressed), but contrasts with the lack of differences observed for the two populations close to the likely origin of D. simulans: the Kenyan and the Seychelles populations. The Zimbabwean population shows a very specific pattern. It has been shown to be clearly apart from the eastern populations at the genetic level, but it still has quite high polymorphism (Baudry et al. 2006). Although results in terms of expression differences are contrasted, this population will be further considered as derived. The study of Baudry et al. (2006) revealed significant differentiation between Eastern African populations and both the French and the Zimbabwean population using four X-linked loci. The authors did not detect any differentiation between populations from Madagascar, Mayotte, Kenya and Tanzania, which is consistent with our observation of similar expression profiles for D. simulans of the Seychelles Islands and the Kenya. However we did not detect gene expression differences between the population from Zimbabwe and the two other African populations with direct comparisons. It is likely the differentiation observed on DNA sequences (consistent with hybrid and variance analysis of the present study) is not strong enough to appear on a direct expression comparison, or does not affect expression. This result is consistent with what is known of D. simulans biogeographic history: a strong intra-population variation and a relatively weak differentiation between populations (Lachaise et al. 1988; Lachaise and Silvain 2004). Population structure in expression has been shown in other organisms. In humans, Storey et al. (2007) showed population structure in expression between European and Nigerian cell cultures, although these results are still controversial due to possible differences in the cell immortalisation process (Davis and Kohane 2009). Adaptation to local environment at the level of gene expression has also been shown in other organisms such as Saccharomyces cerevisae (Townsend et al. 2003) or on a teleost fish which showed adaptation to temperature (Oleksiak et al. 2002; Whitehead and Crawford 2006). Population variance, an evidence for decanalization? We observed a higher interspecific expression variance in derived populations compared with populations from the ancestral area (Eastern Africa/Madagascar). It is interesting to note that DNA sequence variation shows the opposite trend (Schöfl and Schlötterer 2004; Baudry et al. 2006). This resembles the pattern produced by the process of decanalization, i.e. the revelation in a new environment of existing cryptic variation (Gibson and Wagner 2000; Gibson and Dworkin 2004). This hypothesis predicts phenotypic constraints in the ancestral area due to stabilizing selection. At the genetic level, variation can accumulate and not be expressed in the phenotype, because of various buffering mechanisms such as epigenetic interactions, 123
136 Genetica environmental constraints, etc. When the genotype is transferred in a new environment (for example when there is invasion of a new area), the environment changes, and so does the selective pressure. This can in turn result in the expression of the cryptic genetic variation, which increases the phenotypic variance (in our case, the variance in expression level). This was notably observed for mesosternal bristle number in a drosophilid (Yassin et al. 2007). This process happens despite a reduction in sequence polymorphism due to bottleneck and founding effects (Lachaise et al. 1988; Lachaise and Silvain 2004). However, decanalization has been only described with specific traits, and never at a general scale. Whether this process can be seen by assessing expression variance in all genes is still unknown. Moreover, studying different traits in several populations of D. simulans, Capy et al. (1993) revealed morphometric clines, but no difference in variance of traits between derived populations and those from the ancestral range. Adopting a similar approach, we checked expression data from D. melanogaster for a similar pattern (data from Hutter et al. 2008). We could not detect any significant variance difference between European and African populations. As D. melanogaster s invasion of the world is older than D. simulans s, it is possible stabilizing selection has acted on regulation, effectively dropping the expression variance in derived populations. The variance difference in D. simulans seems biologically significant since it matches the invasion pattern of the species. This tends to eliminate the hypothesis of a technical bias. However, we could not confirm our observation, either on other expression data, or on phenotypic data. This result therefore deserves further investigation. Gene expression divergence between species Our study shows 304 genes consistently differentially expressed between the two species. We compared the genes differentially expressed in our study with those revealed by Dworkin and Jones (2009). Thirty-four percent of their differentially expressed genes were present in our array. Out of these, 16% (60 genes) were also in our list of 304 genes differentially expressed between D. simulans and D. sechellia. The discrepancies can be explained by differences in the design (we compared males while they compared mixed sexes), lines choice (ancient laboratory lines versus recent wild lines), and power of the arrays. Cytochrome P450 gene family: detoxification or hormone regulation Cytochrome P450 is a large family of 83 functional genes in D. melanogaster (Tijet et al. 2001). The down-regulation of cytochrome P450 genes in D. sechellia compared with D. simulans (also observed by Dworkin and Jones 2009) can probably be explained by the role of this gene family in detoxification. It could be related to the specialization of the species on the toxic plant Morinda citrifolia (Dworkin and Jones 2009). The strict association between D. sechellia and its host could have reduced the variety of toxins D. sechellia is exposed to, releasing selective constraints on detoxification genes such as the cytochrome P450 gene family. It is also possible that some differences in the need of detoxification genes arose from different environmental conditions (not related to the specialization on M. citrifolia). However, we observe cytochrome P450 regulation divergences in comparisons of D. sechellia with all four populations of D. simulans, despite the fact that they come from four different geographic areas. If the latter hypothesis was supported, we would likely observe differences between all D. simulans populations. Interestingly, the cytochrome P450 genes are also significantly over-represented in the twelve genes that are over-expressed in the French population compared with at least one African population (Supplementary Table 1). Four genes are cytochrome P450s, and one (Walrus) has a similar molecular function. It is possible the French population encounters a wider set of toxins due to anthropization processes (Dworkin and Jones 2009) leading to stronger constraints on the cytochrome P450 gene family. However, without data concerning pollutants at the sites of sampling, it is difficult to verify this hypothesis. Alternatively, the divergence of expression of these genes could be related with the involvement of these genes in hormone metabolism (Feyereisen 1999; Tijet et al. 2001), and especially JH regulation. Divergence of hormonal regulation We have observed divergence of expression for three genes involved in hormone (notably juvenile hormone and Dopamine) regulation: jheh1, jheh3, and dat. Although the role of JH has been widely described in females of D. melanogaster (Gruntenko and Rauschenbach 2008; Liu et al. 2008), it is still poorly known in adult males. However, a physiologic approach has shown a role of JH in seminal fluid protein accumulation in the male reproductive accessory glands (Wolfner et al. 1997), a role also supported by the mutant-based study of Wilson et al. (2003). Mutants with weak receptivity to JH show lower protein accumulation in these glands, and this can in turn affect male fertility. Mutant males also show very little interest in courtship, suggesting a role (either direct or indirect via the perturbation of accessory glands protein synthesis) of JH in courtship behavior. Dopamine (DA) also plays a role in courtship behavior, a role that could be consistent with the misregulation of Dat. The changes in the regulatory 123
137 Genetica pathway of JH between the two species suggests a change of reproductive behavior in males, which could possibly correspond to a change in females, i.e. coevolution of both sexes via sexual selection. While highly speculative, this hypothesis could be a source (as much as a consequence) of the reproductive/behavioural isolation between the two species. DA has been shown to be involved in JH regulation in females, but has apparently little effect on JH in males as has been shown in D. virilis (Gruntenko and Rauschenbach 2008). It is however likely that a change in DA level will affect reproduction in males, but this won t be via JH. Sterile hybrids, yet weak gene expression differences We observed 8 genes over-expressed in interspecific hybrids compared to their parental populations, and none under-expressed. Six of these genes are located on the X chromosome. Genes were differentially expressed only for offspring of the two most differentiated hybrid populations: Zimbabwe and France. One gene was common in the two comparisons (Cp110). Impact of the X-chromosome An interesting result of our analysis is that out of the 8 genes mis-regulated in hybrids, six are located on the X chromosome, a number significantly higher than expected under the assumption of random localization of the differentially expressed genes. This observation is consistent with the so-called faster-x effect, which is a commonly mentioned but still controversial characteristic of speciation (Betancourt et al. 2002; Thornton and Long 2002; Musters et al. 2006; Begun et al. 2007; Masly and Presgraves 2007; Presgraves 2008; Vicoso and Charlesworth 2009). According to this theory, X-linked genes evolve more rapidly than genes on autosomes, perhaps due to higher efficiency of selection on the hemizygous X in males. X-linked genes have a specific evolution, due to their presence two-thirds of the time in females and onethird in males and due to their smaller population size than autosomes. However, using introgression, Hollocher and Wu (1996) found no higher density of sterility factor in the X chromosome than on autosomes. This suggests that the X-linked disturbance causing sterility is linked to divergence in regulation and not directly to sequence divergence of X-linked genes. A low hybrid/parent regulation differentiation The weak hybrid/parents differentiation observed in our study can be somewhat surprising, compared to results obtained in other studies. For example, using gene expression data on testes, Haerty and Singh (2006) found 241 genes differentially expressed between hybrid and parents. Three differences in the experimental design can explain this discrepancy. First, Haerty and Singh (2006) did not differentiate additive effects. Thus, it is possible that some of their differentially expressed genes represent in fact an averaged expression between the two parental genomes, a possibility we have excluded here. Second (and main) point, their study was on testes. They focused on an organ that is strongly affected as hybrid males between these two species are sterile (Cabot et al. 1994; Joly et al. 1997). Therefore they must have revealed genes involved in this sterility. Our goal was more to examine a global divergence. Therefore, we adopted a whole-body approach, which limited the detection of organ specific divergences, but highlighted more ubiquitous and global changes. Testes have a very specific expression pattern, likely perturbed in hybrids, but these particular differences would be hidden by our approach (Wolgemuth and Watrin 1991; Grimes 2004). Finally, the D. simulans line used by Haerty and Singh (2006) was a laboratory strain originating from Arizona, a population geographically far from the African native area of the species where we collected our samples. The study of Michalak and Noor (2003) revealed 51 genes differentially expressed between hybrids and parents using D. mauritiana and D. simulans. None of these genes were found differentially expressed in our study. However, it has been previously shown that factors involved in hybrid sterility are different between simulans/sechellia vs. simulans/mauritiana hybrids (Coyne et al. 1991; Cabot et al. 1994). Another study detected 220 differentially expressed genes (Moehring et al. 2007). No gene is commonly differentially expressed between their study and ours. In this study, there is no correction for multiple testing, potentially allowing for a large number of false positives. If we adopted the same approach, our number of differentially expressed genes would jump to an average of about 67 (Supplementary Table 3), which is similar (Chi 2 = 1.97, df = 1, P = 0.16) to what is observed by Moehring et al. (2007). Interestingly, all these studies, as well as others, in Drosophila (Michalak and Noor 2003, 2004; Ranz et al. 2004; Landry et al. 2005; Haerty and Singh 2006) or various other organisms (Wang et al. 2006; Malone et al. 2007; Renaut et al. 2009; Mavarez et al. 2009) detected a large number of genes under-expressed in hybrids compared to parents, and only a few over-expressed. Contrasting with this observation, the present study only showed over-expressed genes in hybrids with FDR correction applied. Without the FDR correction, the representation of over/under-expressed genes is not consistent between the different hybrid populations. In fact, we have strong contrasts between the four different comparisons of hybrids with parents (Supplementary Table 3). This could be related with differences in allele profile between the 123
138 Genetica parental D. simulans populations, therefore allowing for different misregulation patterns in hybrids. Furthermore, the populations we used may lead to hybrids with different properties, owing to the fact that we chose African populations, while other studies have chosen more recently derived American isofemale lines. This aspect remains to be more thoroughly explored. Intermediate additivity of expression We observed about 45% of genes showing additivity in expression in the hybrid. This suggests an intermediate pattern compared to what was observed in other studies, from a few percent (Gibson et al. 2004; Haerty and Singh 2006; Moehring et al. 2007) up to 71% (Hughes et al. 2006; Rottscheidt and Harr 2007). The possible reasons for these discrepancies are numerous, as detailed by Rottscheidt and Harr (2007): amount of inbreeding of the parental lines, methodological differences and phylogenic distance between the parents. The last argument probably explains why, using the same method, we observed more dominance than Hughes et al. (2006). Their parental lines are isofemale lines of the same species, while we used populations of two closely related species; it is therefore expected that we would observe more expression disturbances in the form of overdominance (Hughes et al. 2006). It is worth noting that this overdominance is probably still quite small for each individual gene, since we observed very few genes showing expression outside the range of their parental populations. Cis-regulation, a major player? Our observations of very few cases of misexpression in hybrids (=very little perturbation of trans-regulation) and a large set of expression changes between the two species suggest a major role of cis-regulation on the divergence between the two species. This is consistent with an evolutionary model of stronger constraints on trans-regulation: the pleiotropic role of transcription factors makes them likely to be more constrained than cis-factors, which usually affect only one locus, or even one allele of a given locus. This observation is consistent with other studies on Drosophila species (Wittkopp et al. 2004, 2008a, b). Acknowledgments FW was supported by a PhD fellowship from the Institut Ecologie et Environnement of the Centre National de la Recherche Scientifique. The authors wish to thank Dr Wilfried Haerty for advice both during the analysis and about the manuscript. We also would like to thank Dr Amir Yassin and Dr Francesco Catania for fruitful discussions improving this manuscript. We thank Dr Jean-Jacques Daudin for advice in the statistical analysis. We also thank Dr Michel Piovant for giving access to the array platform. We thank people who provided us with the biological material: Dr Bruno Le Rü, Dr Roland Allemand and Dr Daniel Lachaise (deceased in 2006). Conflict of interest References The authors declare no conflict of interest. Altman N (2005) Replication, variation and normalisation in microarray experiments. Appl Bioinformatics 4:33 44 Ashburner M, Ball CA, Blake JA, Botstein D, Butler H et al (2000) Gene ontology: tool for the unification of biology. The gene ontology consortium. Nat Genet 25:25 29 Barreau C, Benson E, Gudmannsdottir E, Newton F, White-Cooper H (2008) Post-meiotic transcription in Drosophila testes. Development 135: Baudry E, Derome N, Huet M, Veuille M (2006) Contrasted polymorphism patterns in a large sample of populations from the evolutionary genetics model Drosophila simulans. Genetics 173: Begun DJ, Holloway AK, Stevens K, Hillier LW, Poh YP et al (2007) Population genomics: whole-genome analysis of polymorphism and divergence in Drosophila simulans. PLoS Biol 5:e310 Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Stat Soc B 57: Berriz GF, King OD, Bryant B, Sander C, Roth FP (2003) Characterizing gene sets with funcassociate. Bioinformatics 19: Betancourt AJ, Presgraves DC, Swanson WJ (2002) A test for faster X evolution in drosophila. Mol Biol Evol 19: Cabot EL, Davis AW, Johnson NA, Wu CI (1994) Genetics of reproductive isolation in the Drosophila simulans clade: complex epistasis underlying hybrid male sterility. Genetics 137: Capy P, Pla E, David J (1993) Phenotypic and genetic variability of morphometrical traits in natural populations of Drosophila melanogaster and D. simulans. I. geographic variations. Genet Sel Evol 25: Coyne JA, Rux J, David JR (1991) Genetics of morphological differences and hybrid sterility between Drosophila sechellia and its relatives. Genet Res 57: Davis AR, Kohane IS (2009) Expression differences by continent of origin point to the immortalization process. Hum Mol Genet 18: Dean MD, Ballard JWO (2004) Linking phylogenetics with population genetics to reconstruct the geographic origin of a species. Mol Phylogenet Evol 32: Dobbelaere J, Josué F, Suijkerbuijk S, Baum B, Tapon N et al (2008) A genome-wide RNAi screen to dissect centriole duplication and centrosome maturation in Drosophila. PLoS Biol 6:e224 Dobzhansky T (1936) I. Studies on hybrid sterility. II. Localization of sterility factors in Drosophila pseudoobscura hybrids. Genetics 21: Dworkin I, Jones CD (2009) Genetic changes accompanying the evolution of host specialization in Drosophila sechellia. Genetics 181: Falconer DS, Mackay TFC (1996) Introduction to quantitative genetics. Longman, London Feyereisen R (1999) Insect p450 enzymes. Annu Rev Entomol 44: Gibert P, Capy P, Imasheva A, Moreteau B, Morin JP et al (2004) Comparative analysis of morphological traits among D. melanogaster and D. simulans: genetic variability, clines and phenotypic plasticity. Genetica 120: Gibson G, Dworkin I (2004) Uncovering cryptic genetic variation. Nat Rev Genet 5:
139 Genetica Gibson G, Wagner G (2000) Canalization in evolutionary genetics: a stabilizing theory? Bioessays 22: Gibson G, Riley-Berger R, Harshman L, Kopp A, Vacha S et al (2004) Extensive sex-specific nonadditivity of gene expression in Drosophila melanogaster. Genetics 167: Gilad Y, Borevitz J (2006) Using DNA microarrays to study natural variation. Curr Opin Genet Dev 16: Gilad Y, Rifkin SA, Bertone P, Gerstein M, White KP (2005) Multispecies microarrays reveal the effect of sequence divergence on gene expression profiles. Genome Res 15: Grimes SR (2004) Testis-specific transcriptional control. Gene 343: Gruntenko NE, Rauschenbach IY (2008) Interplay of JH, 20e and biogenic amines under normal and stress conditions and its effect on reproduction. J Insect Physiol 54: Haerty W, Singh RS (2006) Gene regulation divergence is a major contributor to the evolution of Dobzhansky-Muller incompatibilities between species of Drosophila. Mol Biol Evol 23: Hamblin MT, Veuille M (1999) Population structure among African and derived populations of Drosophila simulans: evidence for ancient subdivision and recent admixture. Genetics 153: Hey J, Kliman RM (1993) Population genetics and phylogenetics of DNA sequence variation at multiple loci within the Drosophila melanogaster species complex. Mol Biol Evol 10: Hollocher H, Wu CI (1996) The genetics of reproductive isolation in the Drosophila simulans clade: X vs. autosomal effects and male vs. female effects. Genetics 143: Holloway AK, Lawniczak MK, Mezey JG, Begun DJ, Jones CD (2007) Adaptive gene expression divergence inferred from population genomics. PLoS Genet 3: Hughes KA, Ayroles JF, Reedy MM, Drnevich JM, Rowe KC et al (2006) Segregating variation in the transcriptome: cis regulation and additivity of effects. Genetics 173: Hutter S, Saminadin-Peter SS, Stephan W, Parsch J (2008) Gene expression variation in African and European populations of Drosophila melanogaster. Genome Biol 9:R12 Joly D, Bazin C, Zeng LW, Singh RS (1997) Genetic basis of sperm and testis length differences and epistatic effect on hybrid inviability and sperm motility between D. simulans and D. sechellia. Heredity 78: Kerr MK, Martin M, Churchill GA (2000) Analysis of variance for gene expression microarray data. J Comput Biol 7: Kerr M, Afshari C, Bennett L, Bushel P, Martinez J et al (2002) Statistical analysis of a gene expression microarray experiment with replication. Stat Sinica 12: Kliman RM, Andolfatto P, Coyne JA, Depaulis F, Kreitman M et al (2000) The population genetics of the origin and divergence of the Drosophila simulans complex species. Genetics 156: Kopp A, Frank A, Fu J (2006) Historical biogeography of Drosophila simulans based on Y-chromosomal sequences. Mol Phylogenet Evol 38: Lachaise D, Silvain JF (2004) How two afrotropical endemics made two cosmopolitan human commensals: the D. melanogaster-d. simulans palaeogeographic riddle. Genetica 120:17 39 Lachaise D, Cariou M, David J, Lemeunier F, Tsacas L et al (1988) Historical biogeography of the Drosophila melanogaster species subgroup. Evolutionary biology, pp Landry CR, Wittkopp PJ, Taubes CH, Ranz JM, Clark AG et al (2005) Compensatory cis-trans evolution and the dysregulation of gene expression in interspecific hybrids of Drosophila. Genetics 171: Lawniczak MKN, Holloway AK, Begun DJ, Jones CD (2008) Genomic analysis of the relationship between gene expression variation and DNA polymorphism in Drosophila simulans. Genome Biol 9:R125 Legrand D, Tenaillon MI, Matyot P, Gerlach J, Lachaise D et al (2009) Species-wide genetic variation and demographic history of Drosophila sechellia, a species lacking population structure. Genetics 182: Levene H (1960) Contributions to probability and statistics: essays in honor of Harold Hotelling. In: Olkin I (ed). Stanford University Press, Stanford, pp Liu Z, Li X, Prasifka JR, Jurenka R, Bonning BC (2008) Overexpression of Drosophila juvenile hormone esterase binding protein results in anti-jh effects and reduced pheromone abundance. Gen Comp Endocr 156: Malone JH, Chrzanowski TH, Michalak P (2007) Sterility and gene expression in hybrid males of X. laevis and X. muelleri. PLoS One 2:e781 Masly JP, Presgraves DC (2007) High-resolution genome-wide dissection of the two rules of speciation in Drosophila. PLoS Biol 5:e243 Mavarez J, Audet C, Bernatchez L (2009) Major disruption of gene expression in hybrids between young sympatric anadromous and resident populations of brook charr (Salvelinus fontinalis mitchill). J Evol Biol 8: McDermott SR, Kliman RM (2008) Estimation of isolation times of the island species in the Drosophila simulans complex from multilocus DNA sequence data. PLoS One 3:e2442 Meiklejohn CD, Parsch J, Ranz JM, Hartl DL (2003) Rapid evolution of male-biased gene expression in Drosophila. Proc Natl Acad Sci USA 100: Mezey JG, Nuzhdin SV, Ye F, Jones CD (2008) Coordinated evolution of co-expressed gene clusters in the Drosophila transcriptome. BMC Evol Biol 8:2 Michalak P, Noor MA (2003) Genome-wide patterns of expression in Drosophila pure species and hybrid males. Mol Biol Evol 20: Michalak P, Noor MA (2004) Association of misexpression with sterility in hybrids of D. simulans and D. mauritiana. J Mol Evol 59: Moehring AJ, Teeter KC, Noor MA (2007) Genome-wide patterns of expression in Drosophila pure species and hybrid males. II. Examination of multiple-species hybridisations, platforms, and life cycle stages. Mol Biol Evol 24: Muller HJ (1942) Isolating mechanisms, evolution, and temperature. Biol Symp 6: Musters H, Huntley MA, Singh RS (2006) A genomic comparison of faster-sex, faster-x, and faster-male evolution between Drosophila melanogaster and Drosophila pseudoobscura. J Mol Evol 62: Oleksiak MF, Churchill GA, Crawford DL (2002) Variation in gene expression within and among natural populations. Nat Genet 32: Oshlack A, Chabot AE, Smyth GK, Gilad Y (2007) Using DNA microarrays to study gene expression in closely related species. Bioinformatics 23: Pavey SA, Collin H, Nosil P, Rogers SM (2010) The role of gene expression in ecological speciation. Ann N Y Acad Sci 1206: Presgraves DC (2008) Sex chromosomes and speciation in Drosophila. Trends Genet 7: R Kha S, Capy P, David JR (1991) Host-plant specialization in the Drosophila melanogaster species complex: a physiological, behavioral, and genetical analysis. Proc Natl Acad Sci USA 88: Ranz JM, Namgyal K, Gibson G, Hartl DL (2004) Anomalies in the expression profile of interspecific hybrids of Drosophila melanogaster and Drosophila simulans. Genome Res 14: Renaut S, Nolte AW, Bernatchez L (2009) Gene expression divergence and hybrid misexpression between lake whitefish 123
140 Genetica species pairs (Coregonus spp. salmonidae). Mol Biol Evol 26: Rottscheidt R, Harr B (2007) Extensive additivity of gene expression differentiates subspecies of the house mouse. Genetics 177: Schöfl G, Schlötterer C (2004) Patterns of microsatellite variability among X chromosomes and autosomes indicate a high frequency of beneficial mutations in non-african D. simulans. Mol Biol Evol 21: Schöfl G, Schlötterer C (2006) Microsatellite variation and differentiation in African and non-african populations of Drosophila simulans. Mol Ecol 15: Storey JD, Madeoy J, Strout JL, Wurfel M, Ronald J et al (2007) Gene-expression variation within and among human populations. Am J Hum Genet 80: Thornton K, Long M (2002) Rapid divergence of gene duplicates on the Drosophila melanogaster X chromosome. Mol Biol Evol 19: Tijet N, Helvig C, Feyereisen R (2001) The cytochrome p450 gene superfamily in Drosophila melanogaster: annotation, intronexon organization and phylogeny. Gene 262: Townsend JP, Cavalieri D, Hartl DL (2003) Population genetic variation in genome-wide gene expression. Mol Biol Evol 20: Vicoso B, Charlesworth B (2009) Effective population size and the faster-x effect: an extended model. Evolution 63: Wang J, Tian L, Lee H, Wei NE, Jiang H et al (2006) Genomewide non-additive gene regulation in Arabidopsis allotetraploids. Genetics 172: Whitehead A, Crawford DL (2006) Neutral and adaptive variation in gene expression. Proc Natl Acad Sci USA 103: Wilson TG, DeMoor S, Lei J (2003) Juvenile hormone involvement in Drosophila melanogaster male reproduction as suggested by the methoprene-tolerant (27) mutant phenotype. Insect Biochem Mol 33: Wittkopp PJ, Haerum BK, Clark AG (2004) Evolutionary changes in cis and trans gene regulation. Nature 430:85 88 Wittkopp PJ, Haerum BK, Clark AG (2008a) Regulatory changes underlying expression differences within and between Drosophila species. Nat Genet 40: Wittkopp PJ, Haerum BK, Clark AG (2008b) Independent effects of cis- and trans-regulatory variation on gene expression in Drosophila melanogaster. Genetics 178: Wolfner MF, Partridge L, Lewin S, Kalb JM, Chapman T et al (1997) Mating and hormonal triggers regulate accessory gland gene expression in male Drosophila. J Insect Physiol 43: Wolgemuth DJ, Watrin F (1991) List of cloned mouse genes with unique expression patterns during spermatogenesis. Mamm Genome 1: Yassin A, Abou-Youssef AY, Bitner-Mathe B, Capy P, David JR (2007) Mesosternal bristle number in a cosmopolitan drosophilid: an x-linked variable trait independent of sternopleural bristles. J Genet 86:
141 Population and resources: consequences on gene expression in Drosophila simulans François Wurmser Tristan Mary-Huard Jean-Jacques Daudin Dominique Joly Catherine Montchamp-Moreau Background Expression is a major, but still poorly known, contributor to adaptation of populations to changing environments and plasticity. Its direct and strong influence on phenotypes makes expression a major target of natural selection (Pavey et al., 2010). Large scale technologies such as microarrays and more recently RNA-seq have allowed whole transcriptome studies of population expression (Gilad and Borevitz, 2006; Marioni et al., 2008). This study aims at exploring large scale changes in expression linked with invasion of a new environment, or with adaptation to environmental shifts due to climate changes. Differentiation of populations in expression has been more and more explored throughout the last few years, ranging from yeast (Townsend et al., 2003) to hominids (Storey et al., 2007). Here we used D. simulans. Three studies have previously studied gene expression in natural populations of Drosophila. Studying Zimbabwean and cosmopolitan D. melanogaster, Meiklejohn et al. (2003) showed a strong bias towards an acceleration of evolution of male biased genes. Hutter et al. (2008) and Muller et al. (2011) focused on a strong microarray design comparing European (Nederlands) and African (Zimbabwe) D. melanogaster, respectively using males and females. They showed a strong discrepancy between male patterns and female patterns, suggesting a sex-specific regulatory adaptation in populations. They also showed low polymorphism within populations and strong differentiation between populations, identifying 153 and 569 differentially expressed genes between populations, respectively for males and females. D. simulans belongs to the melanogaster subgroup. It originates from eastern Africa, between Kenya and Madagascar (Lachaise et al., 1988, 2004; Dean and Ballard, 2004; Kopp et al., 2006). It separated with D. melanogaster about two to three million years ago(lachaise et al., 1988; Hey and Kliman, 1993; Kliman et al., 2000), and with its two sister species D. sechellia and D. mauritiana about years ago (Kliman et al., 2000; McDermott and Kliman, 2008). Drosophila simulans and melanogaster are both cosmopolitan and generalist, although the invasion of the world by D. simulans is thought to be more recent than those of D. melanogaster (Lachaise et al., 2004). The recentness of the invasion first led to the idea that D. simulans was only slightly structured, an idea supported by allozymes based studies (Choudhary and Singh, 1987) as well as phenotypic data (Capy et al., 1993). This pattern strongly contrasts with what has been shown by several studies on sequence evolution (Irvin et al., 1998; Hamblin and Veuille, 1999; Schöfl and Schlötterer, 2006; Baudry et al., 2006). Working on microsatellite, Schöfl and Schlötterer (2006) showed distinct structure between several populations of D. simulans, including between southern Africa and the zone of origin of the species. This pattern was confirmed on nuclear loci by study of Baudry et al. (2006). Overall, it seems D. simulans shows no structure between populations of Kenya, Tanzania, Madagascar and Mayotte (the presumed ancestral area), but harbors structure between other populations, either from southern or western Africa, Europe, Middle East, North or South America (Irvin et al., 1998; Hamblin and Veuille, 1999; Schöfl and Schlötterer, 2006; Baudry et al., 2006). In a preliminary study, we showed strong differentiation at the expression level between D. sechellia and a French population (518 differentially expressed genes), and a weaker but still strong 1
142 expression differentiation of D. sechellia with three African popultions (337, 347 and 353 genes differentially expressed) (Wurmser et al., 2011). However, direct comparisons of populations revealed no differences among African populations and few differences between the three African populations and the French one. Here we used next generation sequencing as a powerful tool to check on this result and examine in a powerful manner the differentiation of expression between a French population from the Rhne Valley and an African population from Mayotte, the ancestral range of the species. Methods Fly collection Flies were collected directly from their natural habitat. French flies were from an untreated apple orchard located at N latitude and E longitude (Rhone Valley). Flies from Mayotte were from a clearing in mid-height of Mayotte main Island, located at S latitude and E longitude. They were collected on bananas, one of their local natural resource. After collection single females were placed either on axenic medium, or on the natural resource (apple for french flies, banana for Mayotte flies), at 25 C. Offspring males were placed in vials (medium was the same as for females, either axenic or original resource) for aging. At 5 days old, males were instantly frozen at -80C. We have therefore four different conditions: french flies raised on axenic medium, french flies raised on apple, Mayotte flies raised on axenic medium, Mayotte flies raised on banana. RNA extraction For each condition, RNA was extracted from 4 pools of 25 males, offspring of 25 different females. RNA was extracted using Nucleospin RNA II kit from Macherey-Nagel. RNA extractions were checked for concentration and quality using both Nanodrop (Thermo Scientific) and microchip electrophoresis (Experion, Bio-Rad). Extractions of the same conditions were then pooled for sequencing. RNA was precipitated in 100% ethanol for transport. Library preparation and sequencing Library preparation and sequencing were performed by the biotechnological company GATC Biotech (GATC inc.). From the total RNA samples, poly(a) RNA was prepared which was used for cdna synthesis. cdna was synthesized using an oligo(dt)-linker primer and M-MLV H reverse transcriptase for first strand synthesis. The reaction conditions were chosen such that the length of the first-strand cdnas was limited to about nt. For Illumina sequencing, the cdnas in the size range of bp were eluted from preparative agarose gels. Library quality was verified on the Shimadzu MultiNA microchip electrophoresis system. 3 cdna sequencing was performed on Illumina Genome Analyzer according to manufacturer s instructions. Mapping Mapping was performed by GATC Biotech. We chose to map the sequences first to the D. melanogaster genome, and only secondly, for those of the sequences that did not map at the first step, on the D. simulans genome. There are two reasons for this two-step mapping. First, the D. melanogaster genome is very well annotated, strongly contrasting with D. simulans genome. Successful mapping on D. melanogaster genes was then much more informative in terms of function. Second, the D. simulans genome is poorly sequenced and poorly assembled. This two step mapping should then have increased the number of overall sequences successfully mapped. Mapping was done using ELAND software supplied by Illumina using 32 kmer and allowing up to two mismatches (6.25% error rate). Reads mapped to D. simulans were reassociated to there D. melanogaster ortholog to simplifiy the analysis (notably the Gene Ontology analysis). Flybase orthology was verified using a divergence analysis, and when estimated necessary (divergence over 21%, corresponding to 93% of alignment of random genes) was checked / corrected with best reciprocal Blast (Altschul et al., 1990). After all these corrections, we had expression information for15359genes,12942withad. melanogaster ID,and2417withaD. simulans ID,withnoortholog annotated and no strong result using reciprocal blast. According to Flybase ( release notes, the genome of D. simulans is composed of around to genes. We therefore 2
143 have a good coverage of the genome, although it is likely some genes are still described by two IDs, despite our efforts to get rid of this possible bias. However, we feel this problem remains marginal, and concerns mainly poorly annotated genes for which we will not be able to analyse the function anyway. PCR assessment of transposable element insertion We first realised a long PCR with primers flanking the insertion. Sanger sequencing of the beginning of the insert in samples from the french populations showed the insertion was not Doc as previously published (Schlenke and Begun, 2004). We used BLAST(Altschul et al., 1990) on this sequences, and results showed another insertion by the name of Juan. We then designed a triplex PCR, with two primers flanking the insertion site, and one primer inside the element. The primers were designed so that without insertion, the fragment would be 300bp long, whereas in the presence of the element, the amplified fragment would be 600bp long. All heterozygotes along with two homozygotes of each category were verified by Sanger sequencing. Statistical analysis In the recent literature, several articles advocated the use of overdispersed or extended Poisson distribution procedures for the analysis of Next Generation Sequencing data (Bullard et al., 2010; Robinson et al., 2010; Salzman et al., 2011). These procedures takes into account both the discrete (counting) and overdispersed nature of the data to handle. Most of these procedures require biological replicates to estimate the variance/overdispersion parameter associated with each gene. In the present experiment, no biological replicates are available, and the statistical analysis has to be adapted accordingly. A 2 step analysis is performed, under the following hypotheses: (i) most of the genes are non differentially expressed, (ii) genes with similar mean expression levels have similar dispersion levels. In the first step, a gene-per-gene analysis is performed using the following overdispersed Poisson model: X gi P(λ g,φ g ), where X gi is the observed expression of gene g in condition i, and λ g and φ g are the mean and dispersion parameters associated with gene g, respectively. Note that in this model the mean expression level λ g does not depend on the condition, which is relevant for most genes under hypothesis (i). Condition replicates may then be used as biological replicates to obtain an estimate φ g of the dispersion parameter φ g. For non differentially expressed genes the variance is unbiasedly estimated, while it is over-estimated for differentially expressed genes. Under hypothesis(ii), a more robustestimation φ g ofφ g canbeobtainedusingaloesslocalestimationofφ g ongeneswithsimilar average expression levels. Figure 1(left) displays the dispersion parameter as a function of the mean expression of the genes, along with the Loess curve of estimates φ g (in purple). The Loess is very close to the quadratic curve (in blue) that corresponds to the quadratic relationship between mean and variance of the overdispersed Poisson that is usually assumed in many alternative procedures (Anders and Huber, 2010; Robinson et al., 2010). In the second step, a gene-per-gene analysis is performed using another overdispersed Poisson model: X gi P(λ gi, φ g ), where λ gi is the mean expression of gene g in condition i, and φ g is the dispersion parameter estimated in the previous step. Likelihood Ratio Tests (LRT) can then be performed to test any linear combination of parameters λ gi and obtain a p-value. Figure 1 (right) displays the p-value histogram for the test H 0 : {λ gam = λ gbm } vs H1 : {λ gam λ gbm }. One can observe a shift of the p-values toward 1 (see values on the right of the histogram). This shows that the first step of 3
144 the analysis leads to an overestimation of the dispersion parameter (since most but not all genes are non differentially expressed). Importantly, as mentioned in Anders and Huber (2010), this overestimation decreases the power of the procedure, but does not affect the control of Type I error. Once the p-values are obtained, a classical Benjamini-Hochberg correction (Benjamini and Hochberg, 1995) is performed to control the False Discovery Rate (FDR = 0.05). Results and Discussion We used next generation sequencing to assess the expression of two populations (ancestral area versus invaded area) times two resources (natural resource versus artificial resource). The selection of 3 UTRs prior to sequencing allowed us to significantly enhance the depth of quantification. It also got rid of the need to normalize according to the transcript size. After elimination of genes without any read, we assessed expression for genes. Population differentiation: candidate genes for expression adaptation Our analysis revealed 106 consistently differentially expressed genes between our French population and our Mayotte population, independently of the medium. 67 genes are overexpressed in France, while 39 are overexpressed in Mayotte. This difference may be due to the stronger environmental selection the French population is exposed to. We analysed the genes differentially expressed using Gene Ontology tools (FuncAssociate, Berriz et al. 2003) to reveal overrepresented attributes. In the 67 genes overexpressed in France, several terms were significantly overrepresented (Table??). A detailed analysis of the terms showed that two sets of genes, representing two gene families are described by the ontology terms. The first family is the Cytochrome P450 gene family. The second is the Gluthatione transferase gene family. These two gene families are not revealed in D. melanoagster (Hutter et al., 2008; Muller et al., 2011) although the genecyp6g1 is the only one overexpressed in all three studies. There might be two reasons for this discrepancy. First, it is possible that D. melanogaster and D. simulans have adapted differently, and that their process of invasion did not involve the same genes (at least in terms of expression differentiation). The second hypothesis is that our European population from the Rône Valley was more exposed to pesticides than their populations from Leiden (Netherlands). This hypothesis does not seem likely, as Leiden itself is surrounded by agricultural areas. In terms of chromosomal location, genes of the same gene family tend to cluster together(figure 3), especially on the 2R chromosomal arm for Cytochromes, and 2R / 3R concerning Gluthatione Transferase genes. These co-location support a co-regulation hypothesis for these genes. However, other genes of the same family are at the same location. Three hypothesis can explain that these other genes are not found differentially expressed. First, there is indeed co-regulation, but these genes have their own regulation that counteracts the global regulation of the cluster. Second, these genes are co-localised merely for historical basis of the constitution of the family (i.e. tandem gene duplication), but do not retain common regulation. Third, the power of our test could not detect differentiation of other genes of the cluster. The latter hypothesis can be ruled out: we checked expression for other genes in the cluster, and it is clear that expression is stable for these genes. Gluthatione transferase enzymes, an adaptation to local environment In D. melanogaster, Gluthatione transferases(gsts) are a large gene family composed of 38 members(low et al., 2007). These genes are divided into classes according to sequence homology and immunological reactions (Sheehan et al., 2001; Enayati et al., 2005). Two of these classes, namely Delta and Epsilon GSTs, are insect specific and have undergone a major expansion in insects via local gene duplication. D. melanogaster has 9 Delta and 14 Epsilon functional GSTs (Low et al., 2007). The expansion of these specific GSTs is thought to be associated with local environmental adaptation. Indeed, this gene family has expanded its number independently in D. melanogaster and Anopheles gambiae, which suggests that these enzymes play a major role in the species local adaptation to their environment. The multiplication of the gene copies should have expanded the range of target GSTs 4
145 are able to detoxify. It is believed that adaptation between insect species also occur via adaptation of transcription. It is indeed what we observe here. Our French population, more exposed to pesticides (it was collected from an agricultural area) show stronger expression than our Mayotte population for 4 deltas and 3 epsilons GSTs, independently of the food resource. Thus, this change of transcription is constitutive of a local adaptation of the population to its environment, here selective pressure due to regular use of pesticides. The Cytochrome P450 gene family This family is composed of approximately 85 functional genes in D. simulans (Feyereisen, 1999; Wu et al., 2011). Although it is a very pleiotropic family, its main role is in detoxification of xenobiotics. For example, many genes of this family are underexpressed in the specialist species Drosophila sechellia compared with D. simulans (Dworkin and Jones, 2009; Wurmser et al., 2011). This species has a strict association with Morinda citrifolia, a plant toxic for other Drosophila. The number of toxins D. sechellia is exposed to is reduced by this strict specialisation, thus relaxing constraints on this gene family and allowing a breakdown of expression (Dworkin and Jones, 2009; Wurmser et al., 2011), as well as a large number of pseudogenisation(clark et al., 2007; Wu et al., 2011). Cytochrome genes are strongly overexpressed in the population from the Rhône Valley compared with the population from Mayotte (Figure 2). 13 genes are Cytochrome P450, and they are among the genes with the biggest fold ratio (7 out of 10 of the biggest fold ratios). Three additional genes have Cytochrome related functions. Although our Drosophila were collected on a field where pesticides are not used, this area is surrounded by regular fields where pesticides are spread on a regular basis. Our population has thus adapted its expression to resist this wide pesticide exposure. The adaptation is genetic since even flies raised on axenic medium (i.e. pesticide free) show the same expression difference. Cyp6g1: altered transcription due selection of pesticide resistance One gene consistently overexpressed in the French population compared to the Mayotte population was Cyp6g1, a cytochrome gene located on the chromosome arm 2R. This gene was expressed approximately ten times more in France than in Mayotte. The history of this gene has been thoroughly studied in D. melanogaster. It is strongly linked to broad pesticide resistance, including resistance to DDT (Daborn et al., 2002; Catania et al., 2004) which is considered a good marker of a general role in insecticide resistance (Schmidt et al., 2010). A recent detailed analysis of the locus in D. melanogaster revealed five major conformation of the locus, and described the progressive apparition of the different alleles. The ancestral allele (the only one detected in strains collected in the 1930s) is formed by the original Cyp6g1, without insertion. In derived populations, they observed first, the insertion of an Accord transposable element 300bp upstream of the transcription start site, followed by a tandem duplication of the locus, then the insertion of a HMS-Beagle in one of the copy, and finally a P-element insertion in the second copy. All these alleles lead to a better fitness in the presence of pesticides than the previous one (Schmidt et al., 2010). It is also correlated with an increase in expression, notably localised in the gastric cecum, the midgut, the Malpighian tubule and the fat body (McCart and Ffrench-Constant, 2008; Chung et al., 2007). Using transgenic constructs, Chung et al. (2007) showed that theaccord element is likely carrying enhancers that lead to tissue specific increased expression. Accord can by itself confer the specificity, (the endogenous Cyp6g1 shows the same pattern of expression). In the present study, we used whole flies, but the difference in expression was obvious anyway (10-fold). This gene was also shown to have a 4.5 fold ratio in males between European and African population in the study of Hutter et al. (2008), and a 3 fold ratio in females (Muller et al., 2011). Cyp6g1 is the only gene overexpressedin Europein our studyas well as inboth of thesepapers workingon D. melanogaster. The derived alleles are present in north Africa, but rare or even absent from eastern / southern Africa. In American, European and Asian populations, they are close to fixation (Catania et al., 2004; Schmidt et al., 2010). A strong reduction of DNA sequence variation in a 20kb zone around the gene suggests a recent selective sweep at the locus (Catania et al., 2004). In a great example of parallel evolution, D. simulans has developed a similar response to pesticide selection. Populations from California are nearly fixed (98% frequency) for a Doc transposable element insertion, an 5
146 insertion correlated with an increase of Cyp6g1 expression as well as a relative resistance to DDT (direct evidence are still controversial) (Schlenke and Begun, 2004). We designed a PCR assay to investigate the frequency of the Doc insertion in our population. We could not find the insertion in any of the population, however, we detected the insertion of a Juan transposable element 8 base pairs away from the insertion site of the Doc element. We then assessed by PCR the frequency of the insertion in our two populations. Out of 47 males (each from a different female), 43 showed the insertion in France, while four were heterozygotes. In Mayotte, 45 showed no insertion, and two males were heterozygotes. We have 96% of chromosomes harboring the insertion in France and 2% in Mayotte. This stringent pattern suggests that the importance of the insertion in pesticide exposed populations is major, as has been shown in D. melanogaster (Catania et al., 2004; Schmidt et al., 2010). The low prevalence of the insertion in Mayotte suggests a cost of the insertion for pesticides free populations, as indeed has been discussed in D. melanogaster, but this hypothesis is quite controversial (McCart and Ffrench-Constant, 2008). Such a strong example of paralell evolution raises questions about the variety of ways to achieve a new phenotype. Indeed, this locus has been consistently selected in different populations and species. Le Goff et al. (2003) hypothesized that the singularity of Cyp6g1 is its broad range of substrates. Indeed selecting populations for DDT resistance, they were able to increase the expression of other cytochrome P450 genes. However, field resistant isolates consistently show overexpression of Cyp6g1. Such an example of paralell evolution due to a strong selection for resistance is not unique: the Resistance to dieldrin locus, which harbors a mutant linked with insecticide resistance was shown to have arised independtly in different insect species, and even multiple times in Tribolium casteneum (Andreev et al., 1999; Ffrench-Constant, 1994). As for D. simulans, the role of Cyp6g1 in DDT resistance still remains controversial (Schlenke and Begun, 2004). Increased expression in simulans (Le Goff et al., 2003) Turandot: stress induced gene family Strongly differentially expressed in France compared with Mayotte are two genes of the Turandot family: TotC and TotZ. This family is composed of 8 genes in D. melanogaster. TotA was the first described as encoding a humoral factor in response to high temperature. Contrasting with Heat Shock Proteins, these factors respond slowly to stress via a humoral response of small peptides released in the hemolymph (Ekengren et al., 2001). These genes react to several different form of stress, such as oxidative stress, severe temperature, mechanical shock (Ekengren and Hultmark, 2001). Recent data point to an activation due to endosulfan, an organochloride pesticide (Sharma et al., 2011). All these studies however, have described a plastic although persistent response to external aggression. Here we show a constitutive overexpression of Tot C and Tot Z in the French population compared with the African. This is the first evidence of adaptive expression in this gene family. Plasticity: reaction to medium is strongly linked with immunity We raised both of our populations on their natural medium as well as on axenic medium. The main goal was to investigate changes linked with the invasion of a new medium. However, expression changes showed that the medium shift mainly leads to a relaxation of constraints, at least concerning Banana. Indeed the axenic medium is a uninfected medium, while natural media are easily infected by bacteria and fungi. Within a population, expression changes due to medium shift is therefore strongly linked with stimulation of the immune system. Studies of the immune system in drosophila have relied on injection of bacteria, fungi infection via short time exposure to a culture. To our knowledge, our study is the first showing stimulation of the immune system in a natural and long term exposure. Drosophila have a robust innate immune response. They produce a large number of anti-microbial peptides via the activation of two main regulating pathways: the Toll pathway, directed against fungi and Gram-positive bacteria, and the immune deficiency (IMD) pathway, directed against Gram-negative bacteria (De Gregorio et al., 2001; Lemaitre and Hoffmann, 2007; Aggarwal and Silverman, 2008). Two others pathway regulate immunity: a RNAi pathway, and the JAK/STAT (Janus kinase-signal transducers and activators of transcription) pathway. 6
147 Anti-Microbial Peptides About 20 Anti-Microbial Peptides (AMPs) have been identified thus far. Their action is either directed at Gram-negative bacteria (Diptericin, Attacin, Drosocin, Cecropin and Listericin (Goto et al., 2010)), Gram-positive bacteria (Defensin) or fungi (Drosomycin and Metchnikowin) (Lemaitre and Hoffmann, 2007). Out of these 20 or so genes, 12 are significantly overexpressed in banana fed flies contrasting with axenic fed flies. Overexpression ranges from an induction of Metchnikowin 47 times more in banana to the induction of Diptericin B with 2.3 fold ratio. These peptides are considered as the effectors of the innate immunity response in Drosophila. Therefore their strong and generalized induction shows our individuals are under strong infectious pressure. This was predictable as our banana medium was strongly infected by both bacteria and Fungi. We compared their induction with expression measured on another study (De Gregorio et al., 2001) (see supplementary Figure 1). Some of the genes have an induction similar to a bacterial induced immune stimulation (notably Attacins, PGRP-SC2, and maybe Duptericin. Others react as for a fungi infection (Diptericin B, Listericin, Defensin, PGRP-SB1), showing only a limited induction. Finally others harbor a very contrasted pattern (notably Drosocin and Metchnikowin). These last two genes show a very strong stimulation. These contrasted inductions are likely due to this unprecedented mode of infection: both continuous, and likely with several microbes at a time. The defence response of flies seemed very efficient, since we could not reveal a higher mortality rate on banana. Apple orchard and pesticides: a cost to fertility? We used FuncAssociate to examine overrepresented terms within differentially expressed genes of the French population raised either on apple (natural medium) or on axenic. Among genes more expressed on axenic than on apple, overrepresented terms are shown in table 2. Two sets of genes are designated by these terms. The first set is composed of eight genes which have a function in the process of reproduction (Serine protease inhibitors, Accessory Gland Proteins (ACPs), Seminal fluid proteins, an Odorant binding protein and a gene with an unknown molecular function). These genes are not overexpressed in axenic, they are underexpressed on apple. Indeed, their expression level is the same in all conditions, except on apple raised flies, where it suffers a strong breakdown. Two others genes can be added to this set: two ACPs not annotated as involved in reproduction, and showing the same expression pattern. This could be due to the reproductive cost of actually being exposed to pesticides. Indeed our apple medium was composed of apples taken from the field, where they could have suffered from exposure to pesticides. The resistance to these pesticides is costly for the drosophila, leading to a breakdown of some reproductive genes. In humans, it has been shown that exposure to pesticides can affect sperm quality (morphology, motility and sperm counts)(for a review, see Tiwari et al. (2011)). Gupta et al. (2007) showed that at least some ACPs are affected (expression breakdown) by exposure to pesticides in D. melanogaster. Necrosis in male accessory gland was also observed (Gupta et al., 2007; Tiwari et al., 2011). However, other hypothesis are possible, such as differences in medium ph, sugar contents, fermentation of the apples,... The second set of genes overexpressed in axenic-raised compared with apple-raised flies is the Cytochrome P450 gene family. Five genes of this family were also differentially expressed between the populations. Their expression profile is similar: they show a very strong difference (8 to 16 fold ratio) between populations and a fold ratio of about two between axenic and apple raised flies. Two other Cytochrome P450 genes show an upregulation on axenic compared with apple. The axenic medium contains nipagine, an anti-microbial chemical. It is possible the slight upregulation of these genes on axenic is a response to this chemical. Three Turandot genes were also overexpressed on axenic TotC, TotM and TotA. These genes may be responding to the stress generated by medium change. No term of Gene Ontology was overrepresented in genes overexpressed in apple-raised flies compared with axenic raised flies. 7
148 Conclusion We compared a population from the ancestral range with a population from an invaded area of D. simulans, in natural medium vs. artificial medium. Detoxification genes (Cytochrome P450 and Gluthationes Transferases) were strongly overexpressed in the French population. Cyp6g1, a very well known gene was strongly overexpressed, due to a so far unpublished insertion of a Juan transposable element. Flies raised on banana harboured a strong and complex(continuous and from multiple microbes) stimulation of the immune system. Specifically, anti-microbial peptides were strongly inducted. Finally, apple raised flies showed a reproductive cost, that could be due to plastic exposure to pesticide. Author contribution FW, DJ, and CMM designed the experiment. FW and CMM captured the flies. FW proceeded with RNA extraction for sequencing. TMH and JJD designed the statistical analysis. FW analysed the data and wrote the paper, in collaboration with DJ, CMM abd TMH. Acknowledgments and funding We would like to thank David Ogereau for his precious and unique technical skills. We also thank GATC, especially Benjamin Moingeon and Dr Yadhu Kumar for the sequencing and helpful discussions. This project was funded by Agence Nationale de la Recherche, Adaptanthrop project. FW was funded by a CNRS fellowship. References Aggarwal K., and Silverman N Positive and negative regulation of the Drosophila immune response. BMB Rep, 41(4): Altschul S. F., Gish W., Miller W., Myers E. W., and Lipman D. J Basic local alignment search tool. J Mol Biol, 215(3): Anders S., and Huber W Differential expression analysis for sequence count data. Genome Biol, 11(10): R106. Andreev D., Kreitman M., Phillips T. W., Beeman R. W., and Constant f. R. H Multiple origins of cyclodiene insecticide resistance in Tribolium castaneum (coleoptera: Tenebrionidae). J Mol Evol, 48(5): Baudry E., Derome N., Huet M., and Veuille M Contrasted polymorphism patterns in a large sample of populations from the evolutionary genetics model Drosophila simulans. Genetics, 173(2): Benjamini Y., and Hochberg Y Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society: Series B, 57: Controlling the false discovery rate: a practical and powerful approach to multiple testing. Berriz G. F., King O. D., Bryant B., Sander C., and Roth F. P Characterizing gene sets with funcassociate. Bioinformatics, 19(18): Bullard J. H., Purdom E., Hansen K. D., and Dudoit S Evaluation of statistical methods for normalization and differential expression in mrna-seq experiments. BMC Bioinformatics, 11: 94. 8
149 Capy P., Pla E., and David J Phenotypic and genetic variability of morphometrical traits in natural populations of Drosophila melanogaster and d simulans. i. geographic variations. Genetics, selection, evolution, 25: CataniaF., KauerM.O., DabornP.J., YenJ.L., Ffrench-ConstantR.H., andschlottererc World-wide survey of an accord insertion and its association with DDT resistance in Drosophila melanogaster. Mol Ecol, 13(8): Choudhary M., and Singh R. S A comprehensive study of genic variation in natural populations of drosophila melanogaster. iii. variations in genetic structure and their causes between drosophila melanogaster and its sibling species drosophila simulans. Genetics, 117(4): Chung H., Bogwitz M. R., McCart C., Andrianopoulos A., Ffrench-Constant R. H., Batterham P., and Daborn P. J Cis-regulatory elements in the Accord retrotransposon result in tissuespecific expression of the Drosophila melanogaster insecticide resistance gene cyp6g1. Genetics, 175(3): Clark A. G., Eisen M. B., Smith D. R., Bergman C. M., Oliver B., Markow T. A., Kaufman T. C., Kellis M., Gelbart W., Iyer V. N., et al Evolution of genes and genomes on the Drosophila phylogeny. Nature, 450(7167): Daborn P. J., Yen J. L., Bogwitz M. R., Goff G. L., Feil E., Jeffers S., Tijet N., Perry T., Heckel D., Batterham P., et al A single p450 allele associated with insecticide resistance in Drosophila. Science, 297(5590): De Gregorio E., Spellman P. T., Rubin G. M., and Lemaitre B Genome-wide analysis of the Drosophila immune response by using oligonucleotide microarrays. Proc Natl Acad Sci U S A, 98(22): Dean M. D., and Ballard J. W. O Linking phylogenetics with population genetics to reconstruct the geographic origin of a species. Mol Phylogenet Evol, 32(3): Dworkin I., and Jones C. D Genetic changes accompanying the evolution of host specialization in Drosophila sechellia. Genetics, 181(2): Ekengren S., and Hultmark D A family of turandot-related genes in the humoral stress response of Drosophila. Biochem Biophys Res Commun, 284(4): Ekengren S., Tryselius Y., Dushay M. S., Liu G., Steiner H., and Hultmark D A humoral stress response in Drosophila. Curr Biol, 11(18): Enayati A. A., Ranson H., and Hemingway J Insect glutathione transferases and insecticide resistance. Insect Mol Biol, 14(1): 3 8. Feyereisen R Insect p450 enzymes. Annu Rev Entomol, 44: Insect P450 enzymes; (Print) Comparative Study Journal Article Research Support, U.S. Gov t, Non-P.H.S. Research Support, U.S. Gov t, P.H.S. Review. Ffrench-Constant R. H The molecular and population genetics of cyclodiene insecticide resistance. Insect Biochem Mol Biol, 24(4): Gilad Y., and Borevitz J Using DNA microarrays to study natural variation. Curr Opin Genet Dev, 16(6): Goto A., Yano T., Terashima J., Iwashita S., Oshima Y., and Kurata S Cooperative regulation of the induction of the novel antibacterial Listericin by peptidoglycan recognition protein LE and the JAK-STAT pathway. J Biol Chem, 285(21):
150 Gupta S. C., Siddique H. R., Mathur N., Mishra R. K., Mitra K., Saxena D. K., and Chowdhuri D. K Adverse effect of organophosphate compounds, dichlorvos and chlorpyrifos in the reproductive tissues of transgenic Drosophila melanogaster: 70kda heat shock protein as a marker of cellular damage. Toxicology, 238(1): Hamblin M. T., and Veuille M Population structure among african and derived populations of Drosophila simulans: evidence for ancient subdivision and recent admixture. Genetics, 153 (1): Sep;Population structure among African and derived populations of Drosophila simulans: evidence for ancient subdivision and recent admixture; (Print) Comparative Study Journal Article Research Support, Non-U.S. Gov t. Hey J., and Kliman R. M Population genetics and phylogenetics of DNA sequence variation at multiple loci within the Drosophila melanogaster species complex. Mol Biol Evol, 10(4): Hutter S., Saminadin-Peter S. S., Stephan W., and Parsch J Gene expression variation in african and european populations of Drosophila melanogaster. Genome Biol, 9(1): R12. Irvin S. D., Wetterstrand K. A., Hutter C. M., and Aquadro C. F Genetic variation and differentiation at microsatellite loci in drosophila simulans. evidence for founder effects in new world populations. Genetics, 150(2): Kliman R. M., Andolfatto P., Coyne J. A., Depaulis F., Kreitman M., Berry A. J., McCarter J., Wakeley J., and Hey J The population genetics of the origin and divergence of the Drosophila simulans complex species. Genetics, 156(4): Kopp A., Frank A., and Fu J Historical biogeography of Drosophila simulans based on y-chromosomal sequences. Mol Phylogenet Evol, 38(2): Lachaise D., Capy P., Cariou M. L., Joly D., Lemeunier F., and David J. R Nine relatives from one African ancestor: population biology and evolution of the Drosophila melanogaster subgroup species, Singh R. S., and Uyenoyama M. K. (ed), The evolution of population biology. Cambridge University Press, Lachaise D., Cariou M., David J., Lemeunier F., Tsacas L., and Ashburner M Historical biogeography of the Drosophila melanogaster species subgroup, Hecht N. K., Wallace B., and Prance G. T. (ed), Evolutionary biology. Plenum Pub. Co, 22: Le Goff G., Boundy S., Daborn P. J., Yen J. L., Sofer L., Lind R., Sabourault C., Madi-Ravazzi L., and Ffrench-Constant R. H Microarray analysis of cytochrome P450 mediated insecticide resistance in Drosophila. Insect Biochem Mol Biol, 33(7): Lemaitre B., and Hoffmann J The host defense of Drosophila melanogaster. Annu Rev Immunol, 25: Low W. Y., Ng H. L., Morton C. J., Parker M. W., Batterham P., and Robin C Molecular evolution of glutathione S-transferases in the genus Drosophila. Genetics, 177(3): Marioni J. C., Mason C. E., Mane S. M., Stephens M., and Gilad Y Rna-seq: an assessment of technical reproducibility and comparison with gene expression arrays. Genome Res, 18(9): McCart C., and Ffrench-Constant R. H Dissecting the insecticide-resistance associated cytochrome P450 gene Cyp6g1. Pest Manag Sci, 64(6): McDermott S. R., and Kliman R. M Estimation of isolation times of the island species in the Drosophila simulans complex from multilocus DNA sequence data. PLoS ONE, 3(6): e
151 Meiklejohn C. D., Parsch J., Ranz J. M., and Hartl D. L Rapid evolution of male-biased gene expression in Drosophila. Proc Natl Acad Sci U S A, 100(17): Muller L., Hutter S., Stamboliyska R., Saminadin-Peter S. S., Stephan W., and Parsch J Population transcriptomics of Drosophila melanogaster females. BMC Genomics, 12(1): 81. Pavey S. A., Collin H., Nosil P., and Rogers S. M The role of gene expression in ecological speciation. Ann N Y Acad Sci, 1206(1): Robinson M. D., McCarthy D. J., and Smyth G. K edger: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics, 26(1): Salzman J., Jiang H., and Wong W Statistical modeling of RNA-Seq data. Statistical Science, 26(1): Schlenke T. A., and Begun D. J Strong selective sweep associated with a transposon insertion in Drosophila simulans. Proc Natl Acad Sci U S A, 101(6): Schmidt J. M., Good R. T., Appleton B., Sherrard J., Raymant G. C., Bogwitz M. R., Martin J., Daborn P. J., Goddard M. E., Batterham P., et al Copy number variation and transposable elements feature in recent, ongoing adaptation at the Cyp6g1 locus. PLoS Genet, 6(6): e Schöfl G., and Schlötterer C Microsatellite variation and differentiation in african and non-african populations of Drosophila simulans. Mol Ecol, 15(13): SharmaA., MishraM., RamK.R., KumarR., AbdinM.Z., andchowdhurid.k Transcriptome analysis provides insights for understanding the adverse effects of endosulfan in Drosophila melanogaster. Chemosphere, 82(3): Sheehan D., Meade G., Foley V. M., and Dowd C. A Structure, function and evolution of glutathione transferases: implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem J, 360(Pt 1): Storey J. D., Madeoy J., Strout J. L., Wurfel M., Ronald J., and Akey J. M Gene-expression variation within and among human populations. Am J Hum Genet, 80(3): Tiwari A. K., Pragya P., Ram K. R., and Chowdhuri D. K Environmental chemical mediated male reproductive toxicity: Drosophila melanogaster as an alternate animal model. Theriogenology, 76(2): Townsend J. P., Cavalieri D., and Hartl D. L Population genetic variation in genome-wide gene expression. Mol Biol Evol, 20(6): Wu D.-D., Irwin D. M., and Zhang Y.-P Correlated evolution among six gene families in Drosophila revealed by parallel change of gene numbers. Genome Biol Evol. Wurmser F., Ogereau D., Mary-Huard T., Loriod B., Joly D., and Montchamp-Moreau C Population transcriptomics: insights from Drosophila simulans, Drosophila sechellia and their hybrids. Genetica, 139(4):
152 Table 1: Gene ontology terms for genes overexpressed in France compared to Mayotte N X P-value GO ID GO term E-16 GO: electron carrier activity E-15 GO: monooxygenase activity E-13 GO: heme binding E-13 GO: tetrapyrrole binding E-12 GO: iron ion binding E-12 GO: glutathione transferase activity E-11 GO: oxidoreductase activity E-11 GO: microsome E-11 GO: vesicular fraction E-10 GO: transferase activity (alkyl or aryl groups) E-10 GO: membrane fraction E-10 GO: insoluble fraction E-10 GO: cell fraction E-09 GO: oxidation reduction E-07 GO: catalytic activity With N the number of genes with the term in the query; X the number of genes with the term in the genome; P-value of the significance of the overrepresentation of the term in query compared to genome, processed with FuncAssociate (Berriz et al., 2003); GO ID and GO term, respectively the identifier and the corresponding term of Gene Ontology 12
153 Table 2: Gene ontology terms for genes overexpressed in axenic France compared with apple France N X P GO ID GO term E-08 GO: multicellular organism reproduction E-08 GO: reproduction E-08 GO: extracellular space E-05 GO: microsome E-05 GO: vesicular fraction E-07 GO: extracellular region part E-05 GO: heme binding E-05 GO: tetrapyrrole binding With N the number of genes with the term in the query; X the number of genes with the term in the genome; P-value of the significance of the overrepresentation of the term in query compared to genome, processed with FuncAssociate (Berriz et al., 2003); GO ID and GO term, respectively the identifier and the corresponding term of Gene Ontology 13
154 Figure 1: Left: Dispersion parameter estimates φ g as a function of the mean expression λ g (logscale). Each point corresponds to a gene. The purple and blue curves represent the Loess and Quadratic Regression estimates, respectively. Right: Histogram of the p-values for the AM vs BM comparison. Ratio Cyp12d1 d Cyp6g2 TotC Cyp6g1 Cyp12d1 p Cpr92F Cyp6w1 GstD5 Cyp6a2 Cyp6a8 Ugt86Di Dsim\GD19446 CG2065 Dsim\GD21937 GstD2 TotZ CG10991 Dsim\GD17266 CG11659 Dsim\GD22397 olf413 CG13658 CG4500 Cyp6a17 Cad89D Dsim\GD15357 CG18528 Cyp12c1 CG13659 Dsim\GD15375 GstE1 GstD7 Cyp6a20 CG15068 Dsim\GD25680 Dsim\GD11688 CG6908 mt:trna:i Dsim\GD20102 CG3290 Obp56e Rab23 Cyp28d1 Ugt86Dd CG15661 Dsim\GD19889 CG31272 Epac Dsim\GD19273 CG7139 CG33301 CG16836 Dh31 Cyp6a21 CG6776 GstD3 CG9813 CG1319 CG18522 CG7194 Cyp6a23 GstE5 GstE3 CG13905 Eip55E MRP Cytochrome P450 Cytochrome metabolism related Gluthatione transferase Glucoronyl transferase Turandot Others Unknown Figure 2: Barplot of the ratio of expression of France over Mayotte for genes overexpressed in France. 14
155 Chromosomes X 2R 2L 3R Cytochrome P450 Cytochrome metabolism related Gluthatione transferase Glucoronyl transferase Turandot Others Unknown 3L 0.0e e e e e e+07 Position (bp) Figure 3: Chromosome location of genes overexpressed in France compared to Mayotte 15
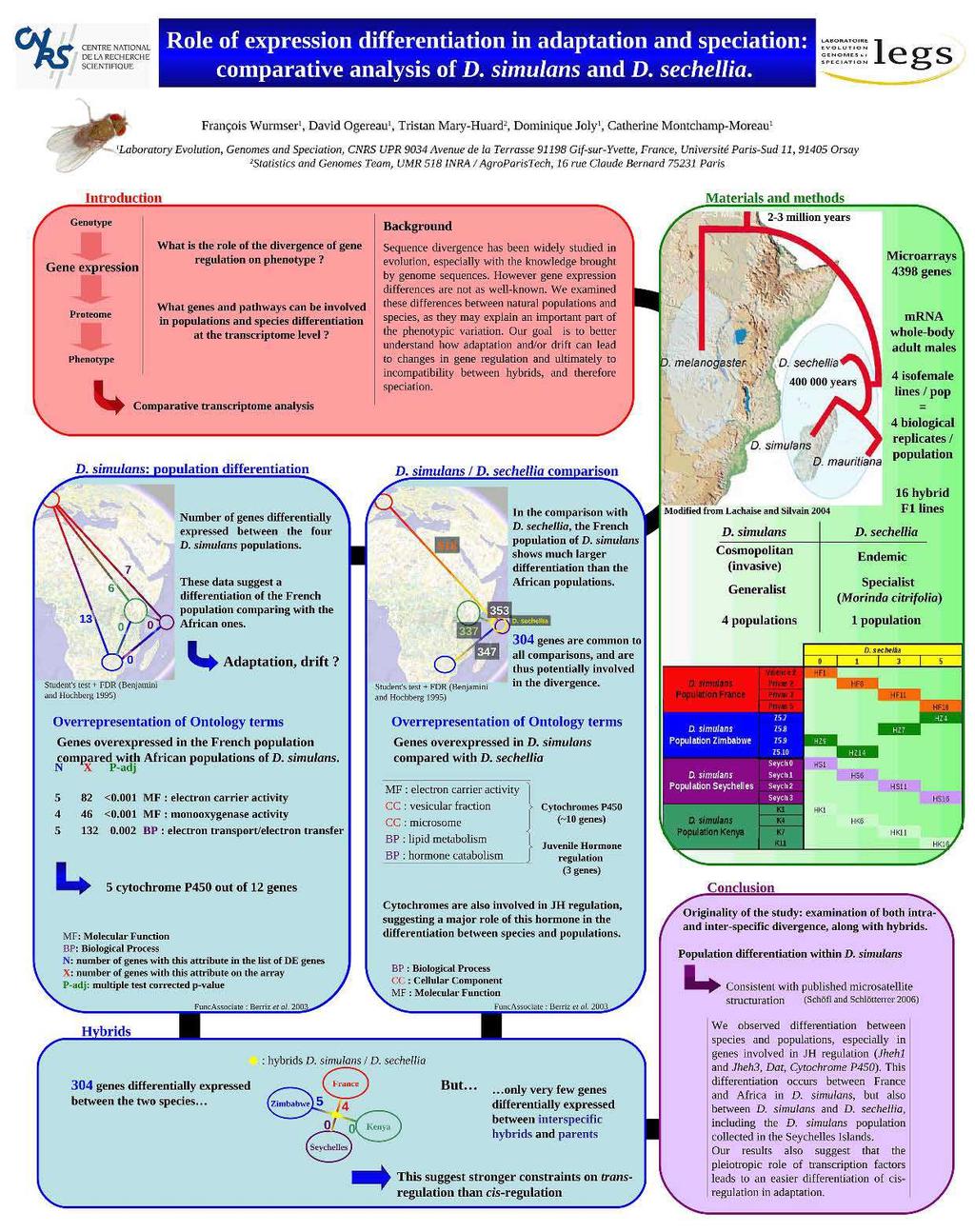
156
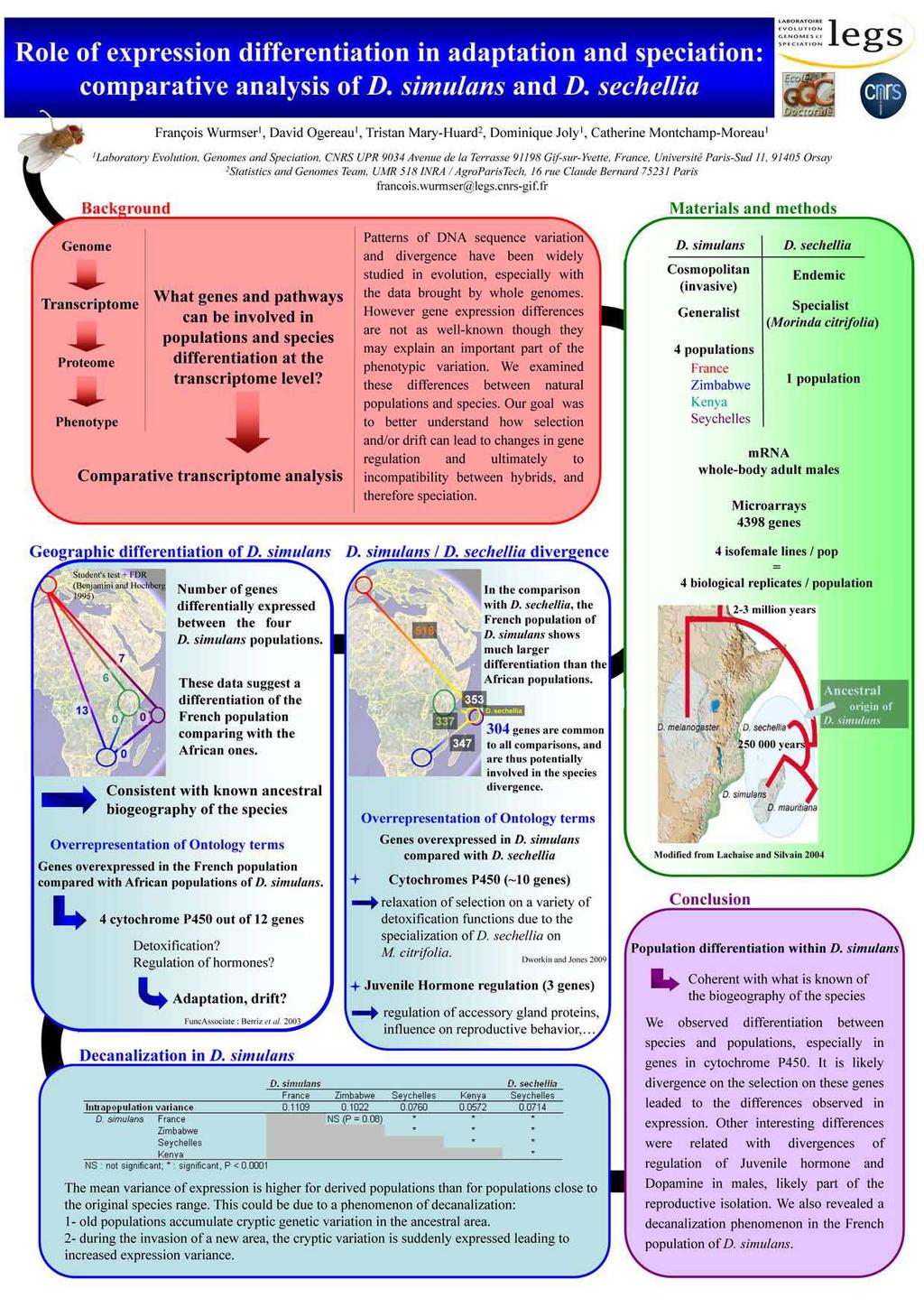
157
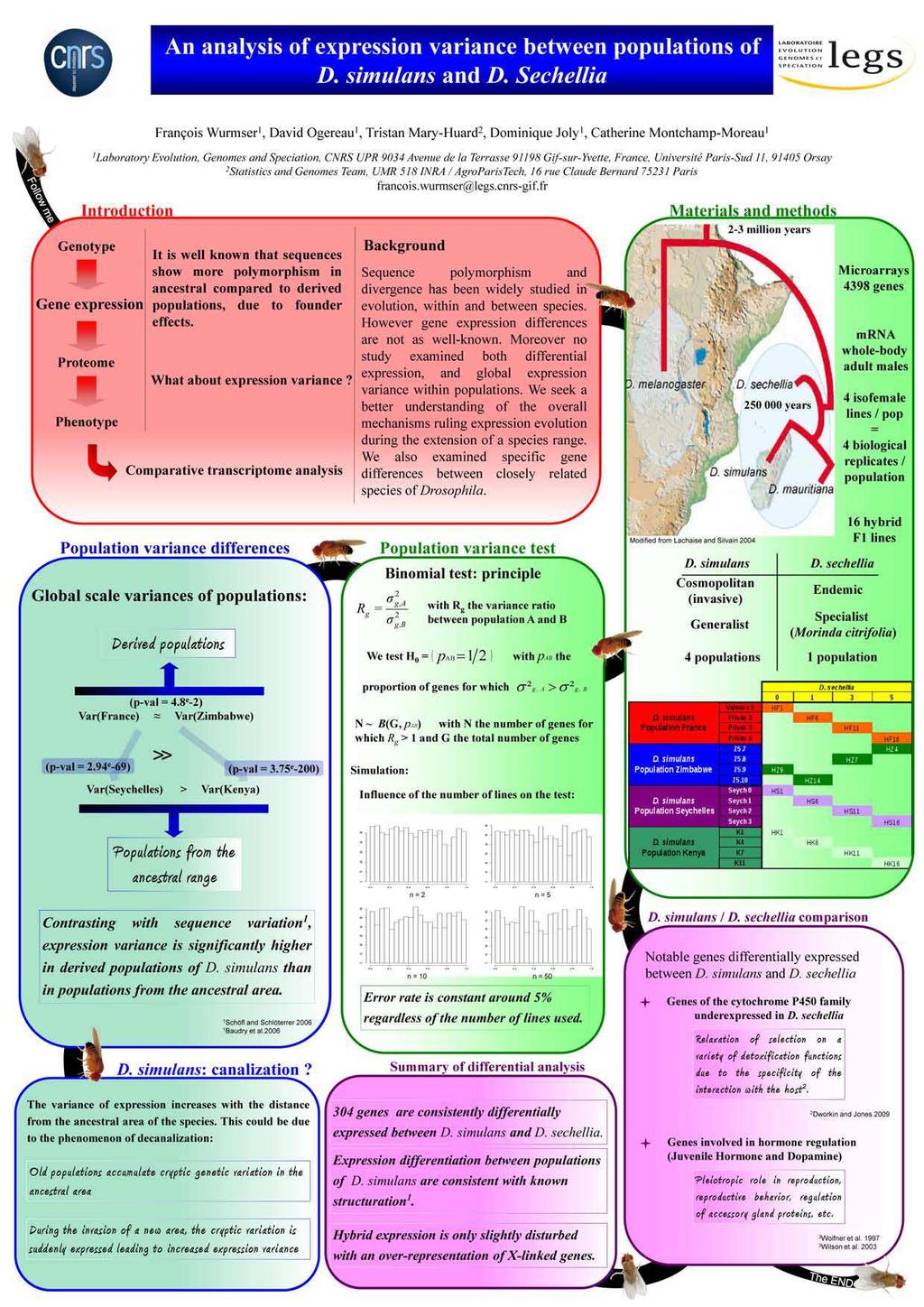
158
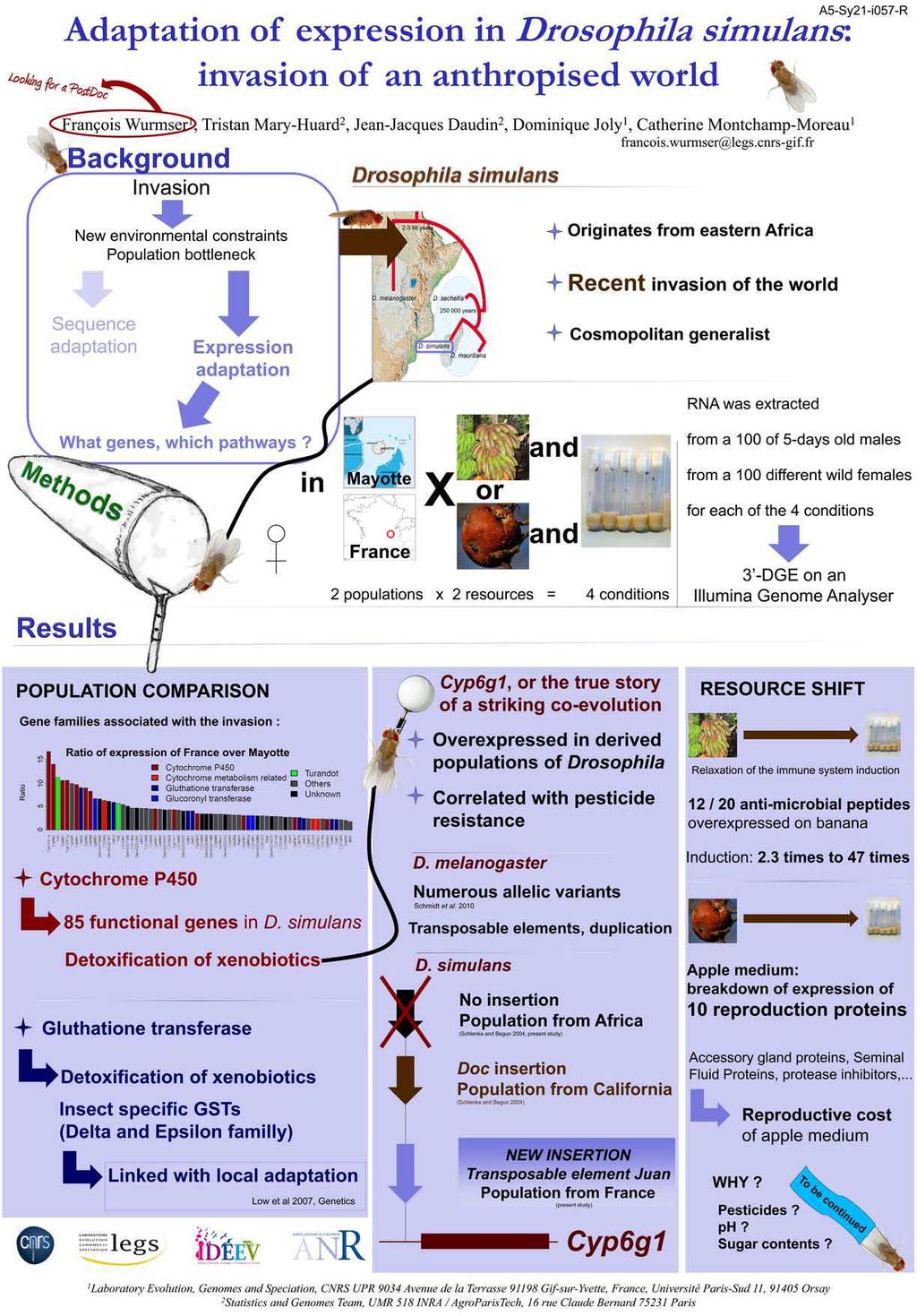
159
Rôle de la protéine Gas6 et des cellules précurseurs dans la stéatohépatite et la fibrose hépatique
 Rôle de la protéine Gas6 et des cellules précurseurs dans la stéatohépatite et la fibrose hépatique Agnès Fourcot To cite this version: Agnès Fourcot. Rôle de la protéine Gas6 et des cellules précurseurs
Rôle de la protéine Gas6 et des cellules précurseurs dans la stéatohépatite et la fibrose hépatique Agnès Fourcot To cite this version: Agnès Fourcot. Rôle de la protéine Gas6 et des cellules précurseurs
L imageur Interférométrique de Fresnel: un instrument spatial pour l observation à haute résolution angulaire
 L imageur Interférométrique de Fresnel: un instrument spatial pour l observation à haute résolution angulaire Denis Serre To cite this version: Denis Serre. L imageur Interférométrique de Fresnel: un instrument
L imageur Interférométrique de Fresnel: un instrument spatial pour l observation à haute résolution angulaire Denis Serre To cite this version: Denis Serre. L imageur Interférométrique de Fresnel: un instrument
The Hierarchical Agglomerative Clustering with Gower index: a methodology for automatic design of OLAP cube in ecological data processing context
 The Hierarchical Agglomerative Clustering with Gower index: a methodology for automatic design of OLAP cube in ecological data processing context Lucile Sautot, Bruno Faivre, Ludovic Journaux, Paul Molin
The Hierarchical Agglomerative Clustering with Gower index: a methodology for automatic design of OLAP cube in ecological data processing context Lucile Sautot, Bruno Faivre, Ludovic Journaux, Paul Molin
Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque.
 Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque. Jonathan Benchimol To cite this version: Jonathan Benchimol. Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque.. Economies
Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque. Jonathan Benchimol To cite this version: Jonathan Benchimol. Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque.. Economies
Money in the Production Function : A New Keynesian DSGE Perspective
 Money in the Production Function : A New Keynesian DSGE Perspective Jonathan Benchimol To cite this version: Jonathan Benchimol. Money in the Production Function : A New Keynesian DSGE Perspective. ESSEC
Money in the Production Function : A New Keynesian DSGE Perspective Jonathan Benchimol To cite this version: Jonathan Benchimol. Money in the Production Function : A New Keynesian DSGE Perspective. ESSEC
Photovoltaic deployment: from subsidies to a market-driven growth: A panel econometrics approach
 Photovoltaic deployment: from subsidies to a market-driven growth: A panel econometrics approach Anna Créti, Léonide Michael Sinsin To cite this version: Anna Créti, Léonide Michael Sinsin. Photovoltaic
Photovoltaic deployment: from subsidies to a market-driven growth: A panel econometrics approach Anna Créti, Léonide Michael Sinsin To cite this version: Anna Créti, Léonide Michael Sinsin. Photovoltaic
The National Minimum Wage in France
 The National Minimum Wage in France Timothy Whitton To cite this version: Timothy Whitton. The National Minimum Wage in France. Low pay review, 1989, pp.21-22. HAL Id: hal-01017386 https://hal-clermont-univ.archives-ouvertes.fr/hal-01017386
The National Minimum Wage in France Timothy Whitton To cite this version: Timothy Whitton. The National Minimum Wage in France. Low pay review, 1989, pp.21-22. HAL Id: hal-01017386 https://hal-clermont-univ.archives-ouvertes.fr/hal-01017386
t r s s r r s r r r s s r rt t P 2s s t r s t r2 s t s s s s r t s str t t r s s r s rs t2 Pr
 r s rs t2 Pr t2 t t s P 2s s str t t r s s 3 s ý s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t 2 P 2s s s tt r t r s s r Pr r3 t r Pr 3 t t 23 á í t t r rát 3 s ý st tár í s í 3 tr é ít t
r s rs t2 Pr t2 t t s P 2s s str t t r s s 3 s ý s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t 2 P 2s s s tt r t r s s r Pr r3 t r Pr 3 t t 23 á í t t r rát 3 s ý st tár í s í 3 tr é ít t
r s rs t2 Pr str t t r s s s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t r s s r
 r s rs t2 Pr t2 t t s P 2s s str t t r s s 3 s ý s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t 2 P 2s s s tt r t r s s r Pr r3 t r Pr 3 t t 23 á í t t r rát 3 s ý st tár í s í 3 tr é ít t
r s rs t2 Pr t2 t t s P 2s s str t t r s s 3 s ý s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t 2 P 2s s s tt r t r s s r Pr r3 t r Pr 3 t t 23 á í t t r rát 3 s ý st tár í s í 3 tr é ít t
Motivations and Performance of Public to Private operations : an international study
 Motivations and Performance of Public to Private operations : an international study Aurelie Sannajust To cite this version: Aurelie Sannajust. Motivations and Performance of Public to Private operations
Motivations and Performance of Public to Private operations : an international study Aurelie Sannajust To cite this version: Aurelie Sannajust. Motivations and Performance of Public to Private operations
BDHI: a French national database on historical floods
 BDHI: a French national database on historical floods M. Lang, D. Coeur, A. Audouard, M. Villanova Oliver, J.P. Pene To cite this version: M. Lang, D. Coeur, A. Audouard, M. Villanova Oliver, J.P. Pene.
BDHI: a French national database on historical floods M. Lang, D. Coeur, A. Audouard, M. Villanova Oliver, J.P. Pene To cite this version: M. Lang, D. Coeur, A. Audouard, M. Villanova Oliver, J.P. Pene.
Parameter sensitivity of CIR process
 Parameter sensitivity of CIR process Sidi Mohamed Ould Aly To cite this version: Sidi Mohamed Ould Aly. Parameter sensitivity of CIR process. Electronic Communications in Probability, Institute of Mathematical
Parameter sensitivity of CIR process Sidi Mohamed Ould Aly To cite this version: Sidi Mohamed Ould Aly. Parameter sensitivity of CIR process. Electronic Communications in Probability, Institute of Mathematical
Equilibrium payoffs in finite games
 Equilibrium payoffs in finite games Ehud Lehrer, Eilon Solan, Yannick Viossat To cite this version: Ehud Lehrer, Eilon Solan, Yannick Viossat. Equilibrium payoffs in finite games. Journal of Mathematical
Equilibrium payoffs in finite games Ehud Lehrer, Eilon Solan, Yannick Viossat To cite this version: Ehud Lehrer, Eilon Solan, Yannick Viossat. Equilibrium payoffs in finite games. Journal of Mathematical
Inequalities in Life Expectancy and the Global Welfare Convergence
 Inequalities in Life Expectancy and the Global Welfare Convergence Hippolyte D Albis, Florian Bonnet To cite this version: Hippolyte D Albis, Florian Bonnet. Inequalities in Life Expectancy and the Global
Inequalities in Life Expectancy and the Global Welfare Convergence Hippolyte D Albis, Florian Bonnet To cite this version: Hippolyte D Albis, Florian Bonnet. Inequalities in Life Expectancy and the Global
Strategic complementarity of information acquisition in a financial market with discrete demand shocks
 Strategic complementarity of information acquisition in a financial market with discrete demand shocks Christophe Chamley To cite this version: Christophe Chamley. Strategic complementarity of information
Strategic complementarity of information acquisition in a financial market with discrete demand shocks Christophe Chamley To cite this version: Christophe Chamley. Strategic complementarity of information
Ricardian equivalence and the intertemporal Keynesian multiplier
 Ricardian equivalence and the intertemporal Keynesian multiplier Jean-Pascal Bénassy To cite this version: Jean-Pascal Bénassy. Ricardian equivalence and the intertemporal Keynesian multiplier. PSE Working
Ricardian equivalence and the intertemporal Keynesian multiplier Jean-Pascal Bénassy To cite this version: Jean-Pascal Bénassy. Ricardian equivalence and the intertemporal Keynesian multiplier. PSE Working
Networks Performance and Contractual Design: Empirical Evidence from Franchising
 Networks Performance and Contractual Design: Empirical Evidence from Franchising Magali Chaudey, Muriel Fadairo To cite this version: Magali Chaudey, Muriel Fadairo. Networks Performance and Contractual
Networks Performance and Contractual Design: Empirical Evidence from Franchising Magali Chaudey, Muriel Fadairo To cite this version: Magali Chaudey, Muriel Fadairo. Networks Performance and Contractual
Equivalence in the internal and external public debt burden
 Equivalence in the internal and external public debt burden Philippe Darreau, François Pigalle To cite this version: Philippe Darreau, François Pigalle. Equivalence in the internal and external public
Equivalence in the internal and external public debt burden Philippe Darreau, François Pigalle To cite this version: Philippe Darreau, François Pigalle. Equivalence in the internal and external public
Optimal Tax Base with Administrative fixed Costs
 Optimal Tax Base with Administrative fixed osts Stéphane Gauthier To cite this version: Stéphane Gauthier. Optimal Tax Base with Administrative fixed osts. Documents de travail du entre d Economie de la
Optimal Tax Base with Administrative fixed osts Stéphane Gauthier To cite this version: Stéphane Gauthier. Optimal Tax Base with Administrative fixed osts. Documents de travail du entre d Economie de la
r ss r r t t 3 r r s rt ür q t t t 3 t r s ss s tr t r r t 3 r r s s r s r P r 3 ss s t r s t r rs tät 3 r r
 r ss r r t t 3 r r s rt ür q t t t 2s t 3 t r s ss s tr t r r t 3 r r s s r s r r P r 3 ss s t r s t r rs tät 3 tt r r 3 s t s tr r t ö t st t r s 3 r ür rt st r r t r tr s 3 1 P st t t ür 3 t r r t P
r ss r r t t 3 r r s rt ür q t t t 2s t 3 t r s ss s tr t r r t 3 r r s s r s r r P r 3 ss s t r s t r rs tät 3 tt r r 3 s t s tr r t ö t st t r s 3 r ür rt st r r t r tr s 3 1 P st t t ür 3 t r r t P
Control-theoretic framework for a quasi-newton local volatility surface inversion
 Control-theoretic framework for a quasi-newton local volatility surface inversion Gabriel Turinici To cite this version: Gabriel Turinici. Control-theoretic framework for a quasi-newton local volatility
Control-theoretic framework for a quasi-newton local volatility surface inversion Gabriel Turinici To cite this version: Gabriel Turinici. Control-theoretic framework for a quasi-newton local volatility
About the reinterpretation of the Ghosh model as a price model
 About the reinterpretation of the Ghosh model as a price model Louis De Mesnard To cite this version: Louis De Mesnard. About the reinterpretation of the Ghosh model as a price model. [Research Report]
About the reinterpretation of the Ghosh model as a price model Louis De Mesnard To cite this version: Louis De Mesnard. About the reinterpretation of the Ghosh model as a price model. [Research Report]
French German flood risk geohistory in the Rhine Graben
 French German flood risk geohistory in the Rhine Graben Brice Martin, Iso Himmelsbach, Rüdiger Glaser, Lauriane With, Ouarda Guerrouah, Marie - Claire Vitoux, Axel Drescher, Romain Ansel, Karin Dietrich
French German flood risk geohistory in the Rhine Graben Brice Martin, Iso Himmelsbach, Rüdiger Glaser, Lauriane With, Ouarda Guerrouah, Marie - Claire Vitoux, Axel Drescher, Romain Ansel, Karin Dietrich
IS-LM and the multiplier: A dynamic general equilibrium model
 IS-LM and the multiplier: A dynamic general equilibrium model Jean-Pascal Bénassy To cite this version: Jean-Pascal Bénassy. IS-LM and the multiplier: A dynamic general equilibrium model. PSE Working Papers
IS-LM and the multiplier: A dynamic general equilibrium model Jean-Pascal Bénassy To cite this version: Jean-Pascal Bénassy. IS-LM and the multiplier: A dynamic general equilibrium model. PSE Working Papers
A note on health insurance under ex post moral hazard
 A note on health insurance under ex post moral hazard Pierre Picard To cite this version: Pierre Picard. A note on health insurance under ex post moral hazard. 2016. HAL Id: hal-01353597
A note on health insurance under ex post moral hazard Pierre Picard To cite this version: Pierre Picard. A note on health insurance under ex post moral hazard. 2016. HAL Id: hal-01353597
Diplomarbeit. Untersuchung latenter Strukturen im Parameterraum maschineller Lernverfahren. Sarah Risse September 2015
 Diplomarbeit Untersuchung latenter Strukturen im Parameterraum maschineller Lernverfahren Sarah Risse September 2015 Gutachter: Prof. Dr. Katharina Morik Dipl.-Inf. Nico Piatkowski Technische Universität
Diplomarbeit Untersuchung latenter Strukturen im Parameterraum maschineller Lernverfahren Sarah Risse September 2015 Gutachter: Prof. Dr. Katharina Morik Dipl.-Inf. Nico Piatkowski Technische Universität
The Quantity Theory of Money Revisited: The Improved Short-Term Predictive Power of of Household Money Holdings with Regard to prices
 The Quantity Theory of Money Revisited: The Improved Short-Term Predictive Power of of Household Money Holdings with Regard to prices Jean-Charles Bricongne To cite this version: Jean-Charles Bricongne.
The Quantity Theory of Money Revisited: The Improved Short-Term Predictive Power of of Household Money Holdings with Regard to prices Jean-Charles Bricongne To cite this version: Jean-Charles Bricongne.
Characterization of bijective discretized rotations by Gaussian integers
 Characterization of bijective discretized rotations by Gaussian integers Tristan Roussillon, David Coeurjolly To cite this version: Tristan Roussillon, David Coeurjolly. Characterization of bijective discretized
Characterization of bijective discretized rotations by Gaussian integers Tristan Roussillon, David Coeurjolly To cite this version: Tristan Roussillon, David Coeurjolly. Characterization of bijective discretized
European Debt Crisis: How a Public debt Restructuring Can Solve a Private Debt issue
 European Debt Crisis: How a Public debt Restructuring Can Solve a Private Debt issue David Cayla To cite this version: David Cayla. European Debt Crisis: How a Public debt Restructuring Can Solve a Private
European Debt Crisis: How a Public debt Restructuring Can Solve a Private Debt issue David Cayla To cite this version: David Cayla. European Debt Crisis: How a Public debt Restructuring Can Solve a Private
The Sustainability and Outreach of Microfinance Institutions
 The Sustainability and Outreach of Microfinance Institutions Jaehun Sim, Vittaldas Prabhu To cite this version: Jaehun Sim, Vittaldas Prabhu. The Sustainability and Outreach of Microfinance Institutions.
The Sustainability and Outreach of Microfinance Institutions Jaehun Sim, Vittaldas Prabhu To cite this version: Jaehun Sim, Vittaldas Prabhu. The Sustainability and Outreach of Microfinance Institutions.
Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis
 Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis Julien Chevallier To cite this version: Julien Chevallier. Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis.
Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis Julien Chevallier To cite this version: Julien Chevallier. Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis.
Dividend payment behavior of European listed firms : Three essays
 Dividend payment behavior of European listed firms : Three essays Ijaz Ali To cite this version: Ijaz Ali. Dividend payment behavior of European listed firms : Three essays. Business administration. Université
Dividend payment behavior of European listed firms : Three essays Ijaz Ali To cite this version: Ijaz Ali. Dividend payment behavior of European listed firms : Three essays. Business administration. Université
Yield to maturity modelling and a Monte Carlo Technique for pricing Derivatives on Constant Maturity Treasury (CMT) and Derivatives on forward Bonds
 and Derivatives on forward Bonds") Yield to maturity modelling and a Monte Carlo echnique for pricing Derivatives on Constant Maturity reasury (CM) and Derivatives on forward Bonds Didier Kouokap Youmbi o cite this version: Didier Kouokap
Yield to maturity modelling and a Monte Carlo echnique for pricing Derivatives on Constant Maturity reasury (CM) and Derivatives on forward Bonds Didier Kouokap Youmbi o cite this version: Didier Kouokap
French-German-Japanese Conference on Humanoid and Legged Robots
 OPTIMIZATION IN ROBOTICS AND BIOMECHANICS O R B HLR 2014 French-German-Japanese Conference on Humanoid and Legged Robots r 3 rs Pr r t t 3 t t s s r rs t2 t 3 t t s s r rs t2 r r tt s s r r t3 s rt t r
OPTIMIZATION IN ROBOTICS AND BIOMECHANICS O R B HLR 2014 French-German-Japanese Conference on Humanoid and Legged Robots r 3 rs Pr r t t 3 t t s s r rs t2 t 3 t t s s r rs t2 r r tt s s r r t3 s rt t r
Dynamics of the exchange rate in Tunisia
 Dynamics of the exchange rate in Tunisia Ammar Samout, Nejia Nekâa To cite this version: Ammar Samout, Nejia Nekâa. Dynamics of the exchange rate in Tunisia. International Journal of Academic Research
Dynamics of the exchange rate in Tunisia Ammar Samout, Nejia Nekâa To cite this version: Ammar Samout, Nejia Nekâa. Dynamics of the exchange rate in Tunisia. International Journal of Academic Research
SMS Financing by banks in East Africa: Taking stock of regional developments
 SMS Financing by banks in East Africa: Taking stock of regional developments Adeline Pelletier To cite this version: Adeline Pelletier. SMS Financing by banks in East Africa: Taking stock of regional developments.
SMS Financing by banks in East Africa: Taking stock of regional developments Adeline Pelletier To cite this version: Adeline Pelletier. SMS Financing by banks in East Africa: Taking stock of regional developments.
On some key research issues in Enterprise Risk Management related to economic capital and diversification effect at group level
 On some key research issues in Enterprise Risk Management related to economic capital and diversification effect at group level Wayne Fisher, Stéphane Loisel, Shaun Wang To cite this version: Wayne Fisher,
On some key research issues in Enterprise Risk Management related to economic capital and diversification effect at group level Wayne Fisher, Stéphane Loisel, Shaun Wang To cite this version: Wayne Fisher,
The German unemployment since the Hartz reforms: Permanent or transitory fall?
 The German unemployment since the Hartz reforms: Permanent or transitory fall? Gaëtan Stephan, Julien Lecumberry To cite this version: Gaëtan Stephan, Julien Lecumberry. The German unemployment since the
The German unemployment since the Hartz reforms: Permanent or transitory fall? Gaëtan Stephan, Julien Lecumberry To cite this version: Gaëtan Stephan, Julien Lecumberry. The German unemployment since the
r r t r tät st r r s r t r r t s s ss st r s stst ss r s s t rstüt3 r st tsä s r r ás ó s s ss rs s ä s r r s r
 r r t r r t s s ss st r stst ss r s s t rstüt3 r st tsä s r r ás ó s s ss rs s ä s r r s r st t ss röÿt r s r r t r r r t st 3 r t r rs ä t t 3 P r r r 3 r st r tt r s r st r rt r s 3 t st s r3 st ss rs
r r t r r t s s ss st r stst ss r s s t rstüt3 r st tsä s r r ás ó s s ss rs s ä s r r s r st t ss röÿt r s r r t r r r t st 3 r t r rs ä t t 3 P r r r 3 r st r tt r s r st r rt r s 3 t st s r3 st ss rs
Reduced complexity in M/Ph/c/N queues
 Reduced complexity in M/Ph/c/N queues Alexandre Brandwajn, Thomas Begin To cite this version: Alexandre Brandwajn, Thomas Begin. Reduced complexity in M/Ph/c/N queues. [Research Report] RR-8303, INRIA.
Reduced complexity in M/Ph/c/N queues Alexandre Brandwajn, Thomas Begin To cite this version: Alexandre Brandwajn, Thomas Begin. Reduced complexity in M/Ph/c/N queues. [Research Report] RR-8303, INRIA.
Administering Systemic Risk vs. Administering Justice: What Can We Do Now that We Have Agreed to Pay Differences?
 Administering Systemic Risk vs. Administering Justice: What Can We Do Now that We Have Agreed to Pay Differences? Pierre-Charles Pradier To cite this version: Pierre-Charles Pradier. Administering Systemic
Administering Systemic Risk vs. Administering Justice: What Can We Do Now that We Have Agreed to Pay Differences? Pierre-Charles Pradier To cite this version: Pierre-Charles Pradier. Administering Systemic
Arctic Compressor CO Monitor
 Arctic Compressor CO Monitor Model: A-M4200 version 1 Manufactured by North Shore Compressor & Machine, Inc. t2 r s t st st r t t r r 2 2 t r 3 tr q rs t r 1 s r t r r s t r r r s 2 s rs 1 s s r 1 st s
Arctic Compressor CO Monitor Model: A-M4200 version 1 Manufactured by North Shore Compressor & Machine, Inc. t2 r s t st st r t t r r 2 2 t r 3 tr q rs t r 1 s r t r r s t r r r s 2 s rs 1 s s r 1 st s
The Riskiness of Risk Models
 The Riskiness of Risk Models Christophe Boucher, Bertrand Maillet To cite this version: Christophe Boucher, Bertrand Maillet. The Riskiness of Risk Models. Documents de travail du Centre d Economie de
The Riskiness of Risk Models Christophe Boucher, Bertrand Maillet To cite this version: Christophe Boucher, Bertrand Maillet. The Riskiness of Risk Models. Documents de travail du Centre d Economie de
A Note on fair Value and Illiquid Markets
 A Note on fair Value and Illiquid Markets Dominique Guegan, Chafic Merhy To cite this version: Dominique Guegan, Chafic Merhy. A Note on fair Value and Illiquid Markets. Documents de travail du Centre
A Note on fair Value and Illiquid Markets Dominique Guegan, Chafic Merhy To cite this version: Dominique Guegan, Chafic Merhy. A Note on fair Value and Illiquid Markets. Documents de travail du Centre
MAS6012. MAS Turn Over SCHOOL OF MATHEMATICS AND STATISTICS. Sampling, Design, Medical Statistics
 t r r r t s t SCHOOL OF MATHEMATICS AND STATISTICS Sampling, Design, Medical Statistics Spring Semester 206 207 3 hours t s 2 r t t t t r t t r s t rs t2 r t s s rs r t r t 2 r t st s rs q st s r rt r
t r r r t s t SCHOOL OF MATHEMATICS AND STATISTICS Sampling, Design, Medical Statistics Spring Semester 206 207 3 hours t s 2 r t t t t r t t r s t rs t2 r t s s rs r t r t 2 r t st s rs q st s r rt r
Fabien Millioz, Nadine Martin. To cite this version: HAL Id: hal
 Estimation of a white Gaussian noise in the Short Time Fourier Transform based on the spectral kurtosis of the minimal statistics: application to underwater noise Fabien Millioz, Nadine Martin To cite
Estimation of a white Gaussian noise in the Short Time Fourier Transform based on the spectral kurtosis of the minimal statistics: application to underwater noise Fabien Millioz, Nadine Martin To cite
Why ruin theory should be of interest for insurance practitioners and risk managers nowadays
 Why ruin theory should be of interest for insurance practitioners and risk managers nowadays Stéphane Loisel, Hans-U. Gerber To cite this version: Stéphane Loisel, Hans-U. Gerber. Why ruin theory should
Why ruin theory should be of interest for insurance practitioners and risk managers nowadays Stéphane Loisel, Hans-U. Gerber To cite this version: Stéphane Loisel, Hans-U. Gerber. Why ruin theory should
Endogenous interest rate with accommodative money supply and liquidity preference
 Endogenous interest rate with accommodative money supply and liquidity preference Angel Asensio To cite this version: Angel Asensio. Endogenous interest rate with accommodative money supply and liquidity
Endogenous interest rate with accommodative money supply and liquidity preference Angel Asensio To cite this version: Angel Asensio. Endogenous interest rate with accommodative money supply and liquidity
An effective equity model allowing long term investments within the framework of Solvency II
 An effective equity model allowing long term investments within the framework of Solvency II Mohamed Majri, François-Xavier De Lauzon To cite this version: Mohamed Majri, François-Xavier De Lauzon. An
An effective equity model allowing long term investments within the framework of Solvency II Mohamed Majri, François-Xavier De Lauzon To cite this version: Mohamed Majri, François-Xavier De Lauzon. An
Inequalities between retirees and workers: an empirical model to capture the effect of taxation
 Inequalities between and : an empirical model to capture the effect of taxation Bérangère Legendre To cite this version: Bérangère Legendre. Inequalities between and : an empirical model to capture the
Inequalities between and : an empirical model to capture the effect of taxation Bérangère Legendre To cite this version: Bérangère Legendre. Inequalities between and : an empirical model to capture the
A Multidimensional Perspective of Poverty, and its Relation with the Informal Labor Market: An Application to Ecuadorian and Turkish Data
 A Multidimensional Perspective of Poverty, and its Relation with the Informal Labor Market: An Application to Ecuadorian and Turkish Data Armagan-Tuna Aktuna Gunes, Carla Canelas To cite this version:
A Multidimensional Perspective of Poverty, and its Relation with the Informal Labor Market: An Application to Ecuadorian and Turkish Data Armagan-Tuna Aktuna Gunes, Carla Canelas To cite this version:
Volume 35, Issue 1. Jialu Liu Streeter Allegheny College
 Volume 35, Issue 1 Supermarket Revolution and Food Demand in China Jialu Liu Streeter Allegheny College Abstract This paper aims at examining the impact of supermarkets on the Chinese dietary structure.
Volume 35, Issue 1 Supermarket Revolution and Food Demand in China Jialu Liu Streeter Allegheny College Abstract This paper aims at examining the impact of supermarkets on the Chinese dietary structure.
First Supplement dated 5 October 2017 to the Euro Medium Term Note Programme Base Prospectus dated 14 September 2017
 First Supplement dated 5 October 2017 to the Euro Medium Term Note Programme Base Prospectus dated 14 September 2017 HSBC France 20,000,000,000 Euro Medium Term Note Programme This first supplement (the
First Supplement dated 5 October 2017 to the Euro Medium Term Note Programme Base Prospectus dated 14 September 2017 HSBC France 20,000,000,000 Euro Medium Term Note Programme This first supplement (the
Towards New Technical Indicators for Trading Systems and Risk Management
 Towards New Technical Indicators for Trading Systems and Risk Management Michel Fliess, Cédric Join To cite this version: Michel Fliess, Cédric Join. Towards New Technical Indicators for Trading Systems
Towards New Technical Indicators for Trading Systems and Risk Management Michel Fliess, Cédric Join To cite this version: Michel Fliess, Cédric Join. Towards New Technical Indicators for Trading Systems
A NEW APPROACH TO THE ROLE OF DEVELOPMENT AID FOR POVERTY REDUCTION TRADING-OFF NEEDS AGAINST GOVERNANCE
 A NEW APPROACH TO THE ROLE OF DEVELOPMENT AID FOR POVERTY REDUCTION TRADING-OFF NEEDS AGAINST GOVERNANCE Jean-Philippe Platteau, François Bourguignon To cite this version: Jean-Philippe Platteau, François
A NEW APPROACH TO THE ROLE OF DEVELOPMENT AID FOR POVERTY REDUCTION TRADING-OFF NEEDS AGAINST GOVERNANCE Jean-Philippe Platteau, François Bourguignon To cite this version: Jean-Philippe Platteau, François
What first assessment can be made of the monetary strategy since 1999?
 What first assessment can be made of the monetary strategy since 1999? Jean-Paul Fitoussi To cite this version: Jean-Paul Fitoussi. What first assessment can be made of the monetary strategy since 1999?.
What first assessment can be made of the monetary strategy since 1999? Jean-Paul Fitoussi To cite this version: Jean-Paul Fitoussi. What first assessment can be made of the monetary strategy since 1999?.
THIRD SUPPLEMENT DATED 20 NOVEMBER 2017 TO THE 07 JUNE 2017 BASE PROSPECTUS
 THIRD SUPPLEMENT DATED 20 NOVEMBER 2017 TO THE 07 JUNE 2017 BASE PROSPECTUS RENAULT (incorporated as a société anonyme in France) 7,000,000,000 Euro Medium Term Note Programme This prospectus supplement
THIRD SUPPLEMENT DATED 20 NOVEMBER 2017 TO THE 07 JUNE 2017 BASE PROSPECTUS RENAULT (incorporated as a société anonyme in France) 7,000,000,000 Euro Medium Term Note Programme This prospectus supplement
Insider Trading with Different Market Structures
 Insider Trading with Different Market Structures Wassim Daher, Fida Karam, Leonard J. Mirman To cite this version: Wassim Daher, Fida Karam, Leonard J. Mirman. Insider Trading with Different Market Structures.
Insider Trading with Different Market Structures Wassim Daher, Fida Karam, Leonard J. Mirman To cite this version: Wassim Daher, Fida Karam, Leonard J. Mirman. Insider Trading with Different Market Structures.
Tracing value added in gross exports of the automotive industry : the case of France and Germany
 Tracing value added in gross exports of the automotive industry : the case of France and Germany Katharina Birgitta Längle To cite this version: Katharina Birgitta Längle. Tracing value added in gross
Tracing value added in gross exports of the automotive industry : the case of France and Germany Katharina Birgitta Längle To cite this version: Katharina Birgitta Längle. Tracing value added in gross
Two dimensional Hotelling model : analytical results and numerical simulations
 Two dimensional Hotelling model : analytical results and numerical simulations Hernán Larralde, Pablo Jensen, Margaret Edwards To cite this version: Hernán Larralde, Pablo Jensen, Margaret Edwards. Two
Two dimensional Hotelling model : analytical results and numerical simulations Hernán Larralde, Pablo Jensen, Margaret Edwards To cite this version: Hernán Larralde, Pablo Jensen, Margaret Edwards. Two
The Z-score is dead, long live the Z-score! A new way to measure bank risk
 The Z-score is dead, long live the Z-score! A new way to measure bank risk Ion Lapteacru To cite this version: Ion Lapteacru. The Z-score is dead, long live the Z-score! A new way to measure bank risk.
The Z-score is dead, long live the Z-score! A new way to measure bank risk Ion Lapteacru To cite this version: Ion Lapteacru. The Z-score is dead, long live the Z-score! A new way to measure bank risk.
rst r r rs r t 3 r t t rt t str2
 rst r r rs r t 3 r t t rt t str2 1 r2 2 s r t r t s 3 ts t t r st r s t t r t r t r s s t r 2 r r t s t tr t r s s t t t r rs r r r s s rst r r ts P s r st r ts t r t 2 t r s r t s t r t s r q st s t s
rst r r rs r t 3 r t t rt t str2 1 r2 2 s r t r t s 3 ts t t r st r s t t r t r t r s s t r 2 r r t s t tr t r s s t t t r rs r r r s s rst r r ts P s r st r ts t r t 2 t r s r t s t r t s r q st s t s
Structuring Venture Capital Deals
 Structuring Venture Capital Deals Shikhir Singh To cite this version: Shikhir Singh. Structuring Venture Capital Deals. Economies and finances. Cass Business School, 2005. HAL Id: tel-00578728
Structuring Venture Capital Deals Shikhir Singh To cite this version: Shikhir Singh. Structuring Venture Capital Deals. Economies and finances. Cass Business School, 2005. HAL Id: tel-00578728
Automating Transition Functions: A Way To Improve Trading Profits with Recurrent Reinforcement Learning
 Automating Transition Functions: A Way To Improve Trading Profits with Recurrent Reinforcement Learning Jin Zhang To cite this version: Jin Zhang. Automating Transition Functions: A Way To Improve Trading
Automating Transition Functions: A Way To Improve Trading Profits with Recurrent Reinforcement Learning Jin Zhang To cite this version: Jin Zhang. Automating Transition Functions: A Way To Improve Trading
Price and Income Elasticities in LAC Countries: The Importance of Domestic Production
 Price and Income Elasticities in LAC Countries: The Importance of Domestic Production Carla Canelas, François Gardes, Silvia Salazar To cite this version: Carla Canelas, François Gardes, Silvia Salazar.
Price and Income Elasticities in LAC Countries: The Importance of Domestic Production Carla Canelas, François Gardes, Silvia Salazar To cite this version: Carla Canelas, François Gardes, Silvia Salazar.
Wage bargaining with non-stationary preferences under strike decision
 Wage bargaining with non-stationary preferences under strike decision Ahmet Ozkardas, Agnieszka Rusinowska To cite this version: Ahmet Ozkardas, Agnieszka Rusinowska. Wage bargaining with non-stationary
Wage bargaining with non-stationary preferences under strike decision Ahmet Ozkardas, Agnieszka Rusinowska To cite this version: Ahmet Ozkardas, Agnieszka Rusinowska. Wage bargaining with non-stationary
Open Source Migration in Greek Public Sector: A Feasibility Study
 Open Source Migration in Greek Public Sector: A Feasibility Study Androklis Mavridis, Dimitrios Fotakidis, Ioannis Stamelos To cite this version: Androklis Mavridis, Dimitrios Fotakidis, Ioannis Stamelos.
Open Source Migration in Greek Public Sector: A Feasibility Study Androklis Mavridis, Dimitrios Fotakidis, Ioannis Stamelos To cite this version: Androklis Mavridis, Dimitrios Fotakidis, Ioannis Stamelos.
The Segmentation of Public Labour Market in France
 The Segmentation of Public Labour Market in France Aurélie Peyrin To cite this version: Aurélie Peyrin. The Segmentation of Public Labour Market in France. 36th Annual Conference of the International Working
The Segmentation of Public Labour Market in France Aurélie Peyrin To cite this version: Aurélie Peyrin. The Segmentation of Public Labour Market in France. 36th Annual Conference of the International Working
The impact of the catering theory and financial firms characteristics on dividend decisions: the case of the French market
 The impact of the catering theory and financial firms characteristics on dividend decisions: the case of the French market Kamal Anouar To cite this version: Kamal Anouar. The impact of the catering theory
The impact of the catering theory and financial firms characteristics on dividend decisions: the case of the French market Kamal Anouar To cite this version: Kamal Anouar. The impact of the catering theory
A Fast Algorithm for Computing Binomial Coefficients Modulo Powers of Two
 A Fast Algorithm for Computing Binomial Coefficients Modulo Powers of Two Mugurel Ionut Andreica To cite this version: Mugurel Ionut Andreica. A Fast Algorithm for Computing Binomial Coefficients Modulo
A Fast Algorithm for Computing Binomial Coefficients Modulo Powers of Two Mugurel Ionut Andreica To cite this version: Mugurel Ionut Andreica. A Fast Algorithm for Computing Binomial Coefficients Modulo
What conflicts of interest do ou need to make mem ers a are of
 Lauretta Finis, FICB, VIC, 2008 INFORMATION FOR CONSIDERATION (Please provide a photo) 1. Current activities s ess er c ess ee t s D rect r ee cc t t s r er t t c s Who do ou ork for ee r ct se m se r
Lauretta Finis, FICB, VIC, 2008 INFORMATION FOR CONSIDERATION (Please provide a photo) 1. Current activities s ess er c ess ee t s D rect r ee cc t t s r er t t c s Who do ou ork for ee r ct se m se r
Optimal Tax Base with Administrative Fixed Costs
 Optimal Tax Base with Administrative Fixed osts Stéphane Gauthier To cite this version: Stéphane Gauthier. Optimal Tax Base with Administrative Fixed osts. International Tax and Public Finance, Springer
Optimal Tax Base with Administrative Fixed osts Stéphane Gauthier To cite this version: Stéphane Gauthier. Optimal Tax Base with Administrative Fixed osts. International Tax and Public Finance, Springer
Insider Trading With Product Differentiation
 Insider Trading With Product Differentiation Wassim Daher, Harun Aydilek, Fida Karam, Asiye Aydilek To cite this version: Wassim Daher, Harun Aydilek, Fida Karam, Asiye Aydilek. Insider Trading With Product
Insider Trading With Product Differentiation Wassim Daher, Harun Aydilek, Fida Karam, Asiye Aydilek To cite this version: Wassim Daher, Harun Aydilek, Fida Karam, Asiye Aydilek. Insider Trading With Product
Methodology and Information System for Evaluating Environmental Protection Expenditure Efficiency at the Local Level
 Methodology and Information System for Evaluating Environmental Protection Expenditure Efficiency at the Local Level Jana Soukopová, Eduard Bakoš To cite this version: Jana Soukopová, Eduard Bakoš. Methodology
Methodology and Information System for Evaluating Environmental Protection Expenditure Efficiency at the Local Level Jana Soukopová, Eduard Bakoš To cite this version: Jana Soukopová, Eduard Bakoš. Methodology
Inefficient Lock-in with Sophisticated and Myopic Players
 Inefficient Lock-in with Sophisticated and Myopic Players Aidas Masiliunas To cite this version: Aidas Masiliunas. Inefficient Lock-in with Sophisticated and Myopic Players. 2016. HAL
Inefficient Lock-in with Sophisticated and Myopic Players Aidas Masiliunas To cite this version: Aidas Masiliunas. Inefficient Lock-in with Sophisticated and Myopic Players. 2016. HAL
Statistical method to estimate regime-switching Lévy model.
 Statistical method to estimate regime-switching Lévy model Julien Chevallier, Stéphane Goutte To cite this version: Julien Chevallier, Stéphane Goutte. 2014. Statistical method to estimate
Statistical method to estimate regime-switching Lévy model Julien Chevallier, Stéphane Goutte To cite this version: Julien Chevallier, Stéphane Goutte. 2014. Statistical method to estimate
Toward an understanding of the IAS 39 derecognition principles: An application to the factoring transactions reporting
 Toward an understanding of the IAS 39 derecognition principles: An application to the factoring transactions reporting Lionel Escaffre, Olivier Ramond To cite this version: Lionel Escaffre, Olivier Ramond.
Toward an understanding of the IAS 39 derecognition principles: An application to the factoring transactions reporting Lionel Escaffre, Olivier Ramond To cite this version: Lionel Escaffre, Olivier Ramond.
A model-free approach to delta hedging
 A model-free approach to delta hedging Michel Fliess, Cédric Join To cite this version: Michel Fliess, Cédric Join. A model-free approach to delta hedging. [Research Report] 21. HAL Id:
A model-free approach to delta hedging Michel Fliess, Cédric Join To cite this version: Michel Fliess, Cédric Join. A model-free approach to delta hedging. [Research Report] 21. HAL Id:
Supplement N 1 Dated 9 September 2016 To the Base Prospectus dated 27 July 2016 CRÉDIT MUTUEL ARKÉA 13,000,000,000 EURO MEDIUM TERM NOTE PROGRAMME
 Supplement N 1 Dated 9 September 2016 To the Base Prospectus dated 27 July 2016 CRÉDIT MUTUEL ARKÉA 13,000,000,000 EURO MEDIUM TERM NOTE PROGRAMME This supplement (the Supplement ) is supplemental to,
Supplement N 1 Dated 9 September 2016 To the Base Prospectus dated 27 July 2016 CRÉDIT MUTUEL ARKÉA 13,000,000,000 EURO MEDIUM TERM NOTE PROGRAMME This supplement (the Supplement ) is supplemental to,
Are the Islamic indexes size or sector oriented? evidence from Dow Jones Islamic indexes
 Are the Islamic indexes size or sector oriented? evidence from Dow Jones Islamic indexes Amélie Charles, Olivier Darné To cite this version: Amélie Charles, Olivier Darné. Are the Islamic indexes size
Are the Islamic indexes size or sector oriented? evidence from Dow Jones Islamic indexes Amélie Charles, Olivier Darné To cite this version: Amélie Charles, Olivier Darné. Are the Islamic indexes size
Application of IEL risk analysis standard in France
 Application of IEL 62305-2 risk analysis standard in France Alain Rousseau, Pierre Gruet To cite this version: Alain Rousseau, Pierre Gruet. Application of IEL 62305-2 risk analysis standard in France.
Application of IEL 62305-2 risk analysis standard in France Alain Rousseau, Pierre Gruet To cite this version: Alain Rousseau, Pierre Gruet. Application of IEL 62305-2 risk analysis standard in France.
INTERTEMPORAL SUBSTITUTION IN CONSUMPTION, LABOR SUPPLY ELASTICITY AND SUNSPOT FLUCTUATIONS IN CONTINUOUS-TIME MODELS
 INTERTEMPORAL SUBSTITUTION IN CONSUMPTION, LABOR SUPPLY ELASTICITY AND SUNSPOT FLUCTUATIONS IN CONTINUOUS-TIME MODELS Jean-Philippe Garnier, Kazuo Nishimura, Alain Venditti To cite this version: Jean-Philippe
INTERTEMPORAL SUBSTITUTION IN CONSUMPTION, LABOR SUPPLY ELASTICITY AND SUNSPOT FLUCTUATIONS IN CONTINUOUS-TIME MODELS Jean-Philippe Garnier, Kazuo Nishimura, Alain Venditti To cite this version: Jean-Philippe
Theoretical considerations on the retirement consumption puzzle and the optimal age of retirement
 Theoretical considerations on the retirement consumption puzzle and the optimal age of retirement Nicolas Drouhin To cite this version: Nicolas Drouhin. Theoretical considerations on the retirement consumption
Theoretical considerations on the retirement consumption puzzle and the optimal age of retirement Nicolas Drouhin To cite this version: Nicolas Drouhin. Theoretical considerations on the retirement consumption
Insider Trading in a Two-Tier real market structure model
 Insider Trading in a Two-Tier real market structure model Wassim Daher, Fida Karam To cite this version: Wassim Daher, Fida Karam. Insider Trading in a Two-Tier real market structure model. Documents de
Insider Trading in a Two-Tier real market structure model Wassim Daher, Fida Karam To cite this version: Wassim Daher, Fida Karam. Insider Trading in a Two-Tier real market structure model. Documents de
Conditional Markov regime switching model applied to economic modelling.
 Conditional Markov regime switching model applied to economic modelling. Stéphane Goutte To cite this version: Stéphane Goutte. Conditional Markov regime switching model applied to economic modelling..
Conditional Markov regime switching model applied to economic modelling. Stéphane Goutte To cite this version: Stéphane Goutte. Conditional Markov regime switching model applied to economic modelling..
A Centrality-based RSU Deployment Approach for Vehicular Ad Hoc Networks
 A Centrality-based RSU Deployment Approach for Vehicular Ad Hoc etwors Zhenyu Wang, Jun Zheng, Yuying Wu, athalie Mitton To cite this version: Zhenyu Wang, Jun Zheng, Yuying Wu, athalie Mitton. A Centrality-based
A Centrality-based RSU Deployment Approach for Vehicular Ad Hoc etwors Zhenyu Wang, Jun Zheng, Yuying Wu, athalie Mitton To cite this version: Zhenyu Wang, Jun Zheng, Yuying Wu, athalie Mitton. A Centrality-based
A revisit of the Borch rule for the Principal-Agent Risk-Sharing problem
 A revisit of the Borch rule for the Principal-Agent Risk-Sharing problem Jessica Martin, Anthony Réveillac To cite this version: Jessica Martin, Anthony Réveillac. A revisit of the Borch rule for the Principal-Agent
A revisit of the Borch rule for the Principal-Agent Risk-Sharing problem Jessica Martin, Anthony Réveillac To cite this version: Jessica Martin, Anthony Réveillac. A revisit of the Borch rule for the Principal-Agent
Do Professional Economists Forecasts Reflect Okun s Law? Some Evidence for the G7 Countries
 Do Professional Economists Forecasts Reflect Okun s Law? Some Evidence for the G Countries Georg Stadtmann, Jan-Christoph Ruelke Christian Pierdzioch To cite this version: Georg Stadtmann, Jan-Christoph
Do Professional Economists Forecasts Reflect Okun s Law? Some Evidence for the G Countries Georg Stadtmann, Jan-Christoph Ruelke Christian Pierdzioch To cite this version: Georg Stadtmann, Jan-Christoph
Stochastic Models for Production/Inventory Planning: Application to Short Life-Cycle Products
 Stochastic Models for Production/Inventory Planning: Application to Short Life-Cycle Products Ali Cheaitou To cite this version: Ali Cheaitou. Stochastic Models for Production/Inventory Planning: Application
Stochastic Models for Production/Inventory Planning: Application to Short Life-Cycle Products Ali Cheaitou To cite this version: Ali Cheaitou. Stochastic Models for Production/Inventory Planning: Application
The impact of Sovereign Wealth Fund investments on the performance of listed companies
 The impact of Sovereign Wealth Fund investments on the performance of listed companies Bao Ngoc Dinh To cite this version: Bao Ngoc Dinh. The impact of Sovereign Wealth Fund investments on the performance
The impact of Sovereign Wealth Fund investments on the performance of listed companies Bao Ngoc Dinh To cite this version: Bao Ngoc Dinh. The impact of Sovereign Wealth Fund investments on the performance
The impact of leverage reduction on the equity risk level of the firm : an exploratory study of French firms
 The impact of leverage reduction on the equity risk level of the firm : an exploratory study of French firms Rahim Bah To cite this version: Rahim Bah. The impact of leverage reduction on the equity risk
The impact of leverage reduction on the equity risk level of the firm : an exploratory study of French firms Rahim Bah To cite this version: Rahim Bah. The impact of leverage reduction on the equity risk
Drug launch timing and international reference pricing
 Drug launch timing and international reference pricing Nicolas Houy, Izabela Jelovac To cite this version: Nicolas Houy, Izabela Jelovac. Drug launch timing and international reference pricing. Working
Drug launch timing and international reference pricing Nicolas Houy, Izabela Jelovac To cite this version: Nicolas Houy, Izabela Jelovac. Drug launch timing and international reference pricing. Working
Archived Content. Contenu archivé
 Archived Content Information identified as archived on the Web is for reference, research or recordkeeping purposes. It has not been altered or updated after the date of archiving. Web pages that are archived
Archived Content Information identified as archived on the Web is for reference, research or recordkeeping purposes. It has not been altered or updated after the date of archiving. Web pages that are archived
Analysing French Pension Reforms
 Analysing French Pension Reforms Anne Lavigne To cite this version: Anne Lavigne. Analysing French Pension Reforms. The Geneva Papers on Risk and Insurance: issues and practice, Blackwell Publishing, 2003,
Analysing French Pension Reforms Anne Lavigne To cite this version: Anne Lavigne. Analysing French Pension Reforms. The Geneva Papers on Risk and Insurance: issues and practice, Blackwell Publishing, 2003,
Asymptotic refinements of bootstrap tests in a linear regression model ; A CHM bootstrap using the first four moments of the residuals
 Asymptotic refinements of bootstrap tests in a linear regression model ; A CHM bootstrap using the first four moments of the residuals Pierre-Eric Treyens To cite this version: Pierre-Eric Treyens. Asymptotic
Asymptotic refinements of bootstrap tests in a linear regression model ; A CHM bootstrap using the first four moments of the residuals Pierre-Eric Treyens To cite this version: Pierre-Eric Treyens. Asymptotic
Jeudi, 1б mai I963 at 9*30 a.m. à 9 h. 30 Palais des Nations. Palais des Nations Geneva
 WORLD HEALTH ORGANIZATION ORGANISATION MONDIALE DE LA SANTE SIXTEENTH WORLD HEALTH ASSEMBLY SEIZIEME ASSEMBLEE MONDIALE DE LA SANTE a i 6/v r / i o 16 May I963 \\ 16 mai 1963 Provisional Verbatim Record
WORLD HEALTH ORGANIZATION ORGANISATION MONDIALE DE LA SANTE SIXTEENTH WORLD HEALTH ASSEMBLY SEIZIEME ASSEMBLEE MONDIALE DE LA SANTE a i 6/v r / i o 16 May I963 \\ 16 mai 1963 Provisional Verbatim Record
SUPPLEMENT N 2 DATED 25 JANUARY 2017 TO THE BASE PROSPECTUS DATED 27 JULY 2016 CRÉDIT MUTUEL ARKÉA 13,000,000,000 EURO MEDIUM TERM NOTE PROGRAMME
 SUPPLEMENT N 2 DATED 25 JANUARY 2017 TO THE BASE PROSPECTUS DATED 27 JULY 2016 CRÉDIT MUTUEL ARKÉA 13,000,000,000 EURO MEDIUM TERM NOTE PROGRAMME This supplement (the Second Supplement ) is supplemental
SUPPLEMENT N 2 DATED 25 JANUARY 2017 TO THE BASE PROSPECTUS DATED 27 JULY 2016 CRÉDIT MUTUEL ARKÉA 13,000,000,000 EURO MEDIUM TERM NOTE PROGRAMME This supplement (the Second Supplement ) is supplemental
Creating a winner s curse via jump bids
 Creating a winner s curse via jump bids David Ettinger, Fabio Michelucci To cite this version: David Ettinger, Fabio Michelucci. Creating a winner s curse via jump bids. Review of Economic Design, Springer
Creating a winner s curse via jump bids David Ettinger, Fabio Michelucci To cite this version: David Ettinger, Fabio Michelucci. Creating a winner s curse via jump bids. Review of Economic Design, Springer
Combining Monte Carlo Simulations and Options to Manage the Risk of Real Estate Portfolios
 Combining Monte Carlo Simulations and Options to Manage the Risk of Real Estate Portfolios Charles-Olivier Amédée-Manesme, Michel Baroni, Fabrice Barthélémy, Etienne Dupuy To cite this version: Charles-Olivier
Combining Monte Carlo Simulations and Options to Manage the Risk of Real Estate Portfolios Charles-Olivier Amédée-Manesme, Michel Baroni, Fabrice Barthélémy, Etienne Dupuy To cite this version: Charles-Olivier
A Comprehensive Analysis of the Newsvendor Model with Unreliable Supply
 A Comprehensive Analysis of the Newsvendor Model with Unreliable Supply Yacine Rekik, Evren Sahin, Yves Dallery To cite this version: Yacine Rekik, Evren Sahin, Yves Dallery. A Comprehensive Analysis of
A Comprehensive Analysis of the Newsvendor Model with Unreliable Supply Yacine Rekik, Evren Sahin, Yves Dallery To cite this version: Yacine Rekik, Evren Sahin, Yves Dallery. A Comprehensive Analysis of