Rôle de la protéine Gas6 et des cellules précurseurs dans la stéatohépatite et la fibrose hépatique
|
|
|
- Oswin McKenzie
- 6 years ago
- Views:
Transcription



1 Rôle de la protéine Gas6 et des cellules précurseurs dans la stéatohépatite et la fibrose hépatique Agnès Fourcot To cite this version: Agnès Fourcot. Rôle de la protéine Gas6 et des cellules précurseurs dans la stéatohépatite et la fibrose hépatique. Médecine humaine et pathologie. Université Paris-Est, Français. <NNT : 2010PEST0040>. <tel > HAL Id: tel Submitted on 16 Mar 2012 HAL is a multi-disciplinary open access archive for the deposit and dissemination of scientific research documents, whether they are published or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
2
3
4 s t èr s t 1t t t s tr t r q t s s s s rés t tr s t è s s s 2 ï s s t s ét s é r ét q t s 1 r s t r s t s ét 1 t t s t r s é é ér t é é ér t é t q à rt r s é t 2t s é é ér t é t q à rt r s ré rs rs s é t t s r q s sté t s t rs trô t ès é t q t rs trô t 12 t s s r s t sé rét s r s s s sté t s sté t s à sté t é t t è s t rs q és s s s ô str ss 12 t é t t r sté t é t t à r s
5 s t èr s é s s r ès ô s s s è s 1 é t t s r q s r té s tr t r t 1 r ss s é t rs s r s t s r ré t r 1 r s t s r s ré t rs r t 2r t s s s t s ô s s r r ô s r ér t r ô s s t r t s r s ô s s t s s t ô s s tr t t r ô s trô t ô s ér t s s ô s 2t s ér t r t t rés t ts ér t r t ô r té s s sté t é t t t r s é t q ô s s s s r s é t q s é ér s r q s 1 1 s r s 11
6 Pr èr rt t 1t t t s
7
8 s s r q s s t r è s té r s s 2s é és sq s t s s réq t s t r s r s r à t ts s t tt s rr s s r ss t r ç s r ét t rés t t r sq r é t r t s r rr s r rt sq s r à s s t st rr s st s s s s s réq t s rr s s t s t 1 ss s t ts s é t t s r s t r t r q s t s r s 2 r é ès r r r t t é ès é t q s s sé èr s ét s r é s r s s2 r ét q tt t é ét q r q t tt t é t 2t r q é ré t t r ré r r t ss r té s és s tr î r t s s é t q q rés t rt é t 2t r t té ré é ér t é rsq t r q t à r t r s t st t st rr s s s t s 2 rt s rt t r s s t ts s r ré t t r ér rt t2 q q s r tér s r r ér t s é t é s 2 t s r tér st q s é t2 q s 1t s r é t 2t r r s s s s s q 3 r t s r s t té s ér r é t 2t s s r s tt ré t t r rr t êtr ré s à rt é t 2t r rr é r r ér t é t 2t s t st t r ré é ér t s rs t r s s ré rs rs s r 1 st rt t r té à s ér r é t 2t s s r s s été é tré 3 r t rr t rés t r ér t t rr s é s ré t s tr t rt rré t tr r s rt ré t t r t t r ér t é t 2t r 3 s t ts tés r r s é t t 2 t sté t é t t q rr s r r t s s r t s s èr t q t s s ré rs rs ss é s 1 2 r st s s ré t t r rr t tr r é t r s ér rt s tr 1 tér rs ré sés s éq tr t q s s ré rs rs é t q s s 2 r st s t s r s sé rèt t r té s r t rr st s r t q s ts t t t q s s é à été q é s r ès tr s r s t s s tr î t ré é ér t é t q s t à és ë tt r té rr t êtr s é t rs s t r t s tr s s s s 2 r st s t s r s t r t s t r q t r ès rs s
9 é t t s r q s rr s ét t s éré rré rs 3 st s r ré t t tr t t s s r q s t rr s q s t s t r t s 1 r r tr t t à r t ét t t rr r s s és s t st s t rs ss t s r s ét s s r s é s s é t s s r q s t r s é t q s t s s s s t s rt r s s t s ré t s t ér t q s t t ès été ét rô r té s t s s s s r ès ss é 1 é t t s r q s P r ét é t t è s s r é t sté t é t t ré t t r t r s s s è s r s r s s é é è r r s q r t ét r rô s s ré rs rs é t q s s s s r ès é t q
10 1 è rt tr t r q
11
12 tr st r s 1 3 s rté rés s t st ré é t r rés t r s r r 3 s t t r t 3 s r s ss r s t s é t t 1 s t é ts ré t ét s é r ét q t s2 t ès r té s q é s s t é t t t r très rt t sq st r èr r é r r s tr s t è s r s2stè st s és s ssè s tés ré é ér t q s t 3 st sé 1 s r t t r q r t s q rs q 3 s r s t r t st st t é s r t q t t t t st ts és r st é 3 t s r s st rr é r s r s t ér t q rt r tr s s r t 3 s t q t té t t s r s rt rt r s q t t s ét t s st rtèr é t q rt rt t rt s 12 é é r r r s r s t ér t st ss ré r s s s s é t q s q r t ér r t é t s és 12 é é rs r




13 t r rés t t t q 3 t 3 r t é r t s és st r sé t t s tés str t r s ré été s s s r s s s é t q s r q st r sé t r tr r t st é té ér ér r s s s rt s q rt t s r t s rtèr t é t q s ss é s à s r s é t 2t s s t r sés tr é s rr é s r s s s ï s ù r s rt t rtér q st s t r é rs tr r t s s s s é t q s r r r s t é t q s s é t q t r s q 3 r s s s rt s s t r és é t q rtèr é t q r t ss 1 2 t q s s é t t s s r sés t r tr r tr r P s rt


14 s s st sé rété r s é t 2t s s s t r é t 2t r r s st té ér ér s s r t s r r t tr s s r s t s r t s r r r à 1tré té s tr é s é t 2t r s r s ré s t st st t é q q s s s t é s à s t t s s r s s é t 2t s t r s tt s t q st t r tt t à s s r r s r tér st q s s ré rs rs t t s q t tr r à ré é ér t rsq s é t 2t s s t térés r é rt é t q tr t tr r s tr é s é t 2t r s tr s s s ï s é t q s t s rt r é r rt r rtèr é t q t r t s s r à 1tré té s tr é s é t 2t r s t t t s s r s s t ss é t q st st t é ér ts t2 s r s q t r ss t tr 1 t s t q és s r s s t s r 2 st st t é r s é t 2t s r sés tr é s r é s r s s s ï s s s s ï s s t é tés r s s t é s tr s tr é s é t 2t r s t s s s ï s s tr s ss r st t é tr 1tr r sé r s è s t

15 s s é st 2 r té s str t r r t t 2s r s 2 s 2 s r té 2 s s s ét é s s s t t s s ss t t r r s és s s s s ï s s tr t s s t r s rés t s r é t ïét q s t2 2é ï s r t q s t r t 2 ï 2 2t s t t r r r s s s t t t s s t é s t str t t é t r 1tr s t s s ss t s t r t s s s ét é s t s é t 2t s r rés t t rt s s ï é t q s ér ts t2 s r s rés ts s t s ss té rès s 2 2t 2 2t t r r t r r s é t 2t s r sés tr é s é t 2t r s st t t r 2 é t q t r rés t t s s s s t s s é t é s r sé s ô s s ï t t tr s ss r t ô r r t tré s é s t r t s t sé rét r té s s2 t ét sé s s s t ô s s ï sé rét t s s st t ts s r s s r ts ét 1 t s t ô r s r tr r t à rt s s ér é s s é t 2t s t é r té r ér t s s rt ér t r tt t ré é ér t rès
16 s s rt é t 2t r s s r s s t s s é t é s q st t t r s 1 r s é t q s s t és s s s s rt s r s 1 r t rs é t q s é q s s r s r rés t t r s s é t q s s s ét é s s t s t sé s s s ss r t st t t à s s é t q s s r é t2 q s t s rés t t r ét é s r ts 2t s q s q ét ss t s t ts s é t 2t s t s s s ï s r r st r s té s s rr t ér r s ré rs rs é t ïét q s s s s t s 1 st s s tt tt s q s t rét r s s sé rèt t r rt s r té s tr 1tr r s s ss s s tr s é t 2t s t s s t é s r s s é s tr s s s ï t s é t 2t s s s ét é s t s tr s r r 2 r st s q 1 r t α t t2 s r ss α r èr t t sé rèt t s 2t s β t s r té s tr 1tr r s s s t ss s s rés t tr s t è s t tr s s 2 2t s t t rô s ré t t r t ré r t s s rés t tr s t è s rô s s rés t tr s t è s st t r s é é ts étr rs t 1 r r à r s r s r ts s t è s ss és à s é s r tt t r ss s s r té s 1 r st t té t 1 st 1 ss s ss st 1 r é r t t s s s éé s t s s tr î str t t s q ss st 1 r é q r s s rés t tr s t è s r ss s s s r t q s t s r s s s s t é s 1 r t t é t s 1 ss s s s t é s r t r s s s ï s t r rés t t r s s é t q s s s t t s ss st t t t é ér t s tr s t é s s r s r s st stré t é r r s r tt t r ss s tr ts t s r é s tr s é t 2t s t s s s ï s s t é s
17 s s é t q s t r t s 2t s é s t rt s s ét s s r té s t s r r t t s t r s r t s rs r ss r 2 é t q s t rô rés t t s t è s s ss t t s t s s2 t ét s r s 2t s t r s t s q t r t r té 2t s tr t t t rés t t s t è s st s r à s s r t q s s s r s t s r s rés ts sés s èr s s s ï s s r rés t t r s s é t q s t s s r s r s s s r t té 2t r s ér t s r t r s s s é t s rt s étr èr s rr t r s s s é t s ts t è s t s s é t q s rès r t s r é r s r 2t s é é r r 1 ré t rs rés ts à s r s s ré t rs s rs t 1 s t 2 sér s t t s s r t s s r t q s é ré s t r é s sé rèt t s 2t s t r s t s q t α t r ér α α t r r s s t r α s q β tr s r r t t r β t s t s r r q rés t t t è r s s t é s s q t t r s s t s s r t q s rés t s s t ér é s ss s t sé s t r s s tr r s s s s rt s s r rés t t s s s s r s s t r t r t t r s ré s sé s à t r r t tr t r s t è s s s r t s s t é s s sé rèt t st t t t t β q ré t r t t ér t r s

18 s s r r r ré t t ré rt t s ér t s t s r s é t q s rès s s 2 ï s s s 2 ï s rés t s r rés t t à s s é t q s r sq s s 2 2t s t r r s 2 2t s t s 2 2t s q r rés t t r s t t t s s 2 ï s é t q s s s s s ï s s 2 2t s s t t t étr t s t è s rés tés r s s t é s s r t s r t q s rés t s s 2 2t s t é t r s t ts s é t 2t s s s ét é s t tr 1tr r s 2 2t s t r r s t s s 2t t 1 q s rés t s s s s ï s é t q s t t é r t s s s t r P t s q ssè t s r s t t s t rs 2t t 1 q s t t r rt t s s r s s 2 ï s q s r t r s t s2 t ét s r r s s 2t s γ t r ér γ t α s s 1 r t à r s r s ré t rs t t rs 1 q r ss t ér ts s 1 r és r s s téré s té s 1 P t s ré t rs t rs 1 q r ss t s t è s s rés té r r 2t t 1 q s s rés t éq r tr s s 1 t t rs

19 s s t t rs rés tés r s s s t té 2t t 1 q s 1 r 1 rt t s r s 2t 2t q s ér t é t rs q tr î t t s s s s r t 2t s t r t é t r 1 st t s s r rés t γ été tré q s s rés t s t t té 2t t 1 q s é é q s s r t s q s é s s r s s s tt s é té s t s s s 2 2t s st t t s s 2 ï s t s t sés 2 2t s t s t t s r s 2 2t s t s t2 t t 1 r t 1 t2 t 2t t 1 q s 1 t2 s rés t t ré rt r r é ré t rs à î s αβ q r ss t s t è s rés tés r ss ss s r rt st q st rs s r é s s r t à s 2 2t s ï s t s s s 2t s r t s s r s r rs q sé rèt s 2t s r t r s r r r t t s 2 2t s ï s rs s é t2 s 2 2t s 1 r s ér ts s 2t s q é s s tt r t t té r s 2 2t s t s s t st t és r s s t s 2 2t s à î γδ r s t r r 1 r t s rq rs s t ré rt r ré t rs très r str t r îtr s 2 s rés tés r é ss ss q 1 r é à é é s r s s t é s s s r t s é t 2t s s s t s ts s q rt t rs s r s t t r rés t r s 2 2t s tr é t q s s s r tér s t r r t r t

20 s s ss 2t s s s 1 r s s t s r à é s 2t s s t ss t2 γ t α q t2 s r ç r s t êtr é r rs tr s ré t rs 1 r és r s s t q rt t à r t t ré s r r t t r 1 s s s t trô é s r s s s s ss q s s t r é s ss t r t 2 s 1 r t ré rt r ré t rs très r str t t r ss t s t è s s t 1t ss é r t è t r st r s t êtr é t s s s ss q s t s ré t rs à î s αβ γδ r t t s t s rt s î s s tr s 2 2t s t s 1 r t s rq rs t s t r t st t és s à ré t rs 1 r ts à î γδ s r t ré rt r r é r îtr r té t è s s r té s str ss t s t è s r té q s r r rés t à s 2 2t s tr é t q s q t s r rés r r s r r s ré rt t s ér t s ss s 2 2t s s t s r t t t r rés t t r rt 2 2t s t s s 1 rt ts
21 s s s 2 2t s r rés t t r s 2 2t s é t q s s t t s s s r r s st r r rés t t s t s tés r s s r q r t r ét s st r 2 2t s t s t st très ér t s 2 2t s r ts r s s 2t t 1 q s t q r rés t t q s 2 2t s r ts s t très t s s t r rés t t à s 1 r té s 2 2t s rés ts s r s 2 2t s t s r rt tr s 1 r s t s 2t t 1 q s st rsé r
22 s t s s t s st r rt t rr é q r ç t s rt r é tr ts t sés t 1 q s t r t s ét s é r ét q s2 t ès r s s r té s t é s r s tr s ts t 1 q s t t 1 st é t r st s rs t s t rés r sq à s t rés r à s t t rés r s t ét 1 r t r è rés r r s rès é s é s s r é à rr t ét s é r ét q rô tr s ét s s s t s s r s q st t t s rés r s é r ét q s r s s tr ts s t s r és t st ss t s r t s rt t rr t r t t s st rr r ét q rt t r ré t é r ét q s st s é r ét q s 1 st t s s r 2 è r 3 êtr r s s t tr 2 ér s s rs s r s s 2 è s tr r s q t tés s s s s à q rés r t à r t t t à rs q s tr 2 ér s s t r t s t és s t ss 1 t s ét s s s rô ss t s é st s s é ss r à rt s t s r s ré 2 é t t s s st s s r 2 è 2 ès t èr t s s s s s r s 2 2s rés 1 ès s ss é à st é é 2 è tr s r s s r s é ès ét s s s t r t é t s ét s q t t s s r s t stér s st s t r q s t ss és s r2 t s t2 r t à rt r stér t s tr 2 ér s q s t s t sé rété s s s t tr s r é s t s t2 r t 2t s s s t2 r t r t ss r r s s s s t ss s stér t t ù st ré rs r s2 t ès s s r s 2 s s é t é r s s r s tés r s t tr s r és tr 2 ér s r 2 êtr st és st ès s é ss té s tr 2 ér s t êtr é r és r 12 t tt
23 s t s 12 t s s r s t r s t r r t s s r 12s s t s r s s β 12 t t r st q é s 12 t s s r s q r t é r à β 12 t t r s st à r r r r r ss t s î s t q s s s tés ét2 q t s s r r s ét q s r s r r s str t é r ét q r s t ss s 1tr é t q s rt r rs û ré tr r tr r s 2 s s tr r 12 q s β 12 t t r st ré é r P r t t 2 tr s r s tr t r t t 2 q P β 12 t s s r 12s s r t ét s s s à très î s r s s s r s ét 2 r és s s r 12 q s s r st ï s r st s t r 1 s r st 2 s t s ér és s s r s à r s s s r s s t s ts s s t 1 q s t q t t s β 12 t ss ré r s r 12s s st r r r s s r s à très î t êtr 12 és s s t r s s t êtr r r s s s r 12s s t r rs r r 12 t s t r s s r 12 q s à î é érés r ω 12 t r s s s r s s t é t 12 és r s r 12s s s s r s t é t êtr 12 és r ω 12 t s s r s s r s 32 s P s 2 r 12 r s s r s s t rés t s t rés q é s s t r ét s s s r s q t té rt t s r 12 q s t êtr r és s s r r r ss s s és té t èt s t ss é à s tés 12 t t r r t ω 12 t s r s s très rt t tér t s tr s ét s ét s é t q s s 1 ès s r s tés r s é t 2t s t ss sé rét s t β 12 t t r r 12s t s2 t ès s r s t tr r à t s q s s s é t 2t s t s 1 r s t r s s2 t ét s r rt s r té s s t s s r s sé rétés s q r tt t é s t s r ss s s é Pr té s s t q s s2 t ét s r s r té s s t q s t s q s r té s t s t rs t t s r té s st
24 s t s r r t t r ss t q r ss s t q é 1 r té s s s t s rt tr s rt s é ts t sés 2 r s r é s s t rs és à t t s2 t ès 1 s t é t q s t r r è t r r t r t r t s t rs t t t té é t t t s r t é ss t s s s r s t ss s2 t ès s è s r té s tr s rt r s r s stér ï s t rt à s2 t ès s r té s t q é s s tr s rt s s P s rs t s r s s t ss ré s é t q t sé rét s r t t r q st r s s s ts r és r r ss t rô s à s s r t t r s st é t tr s r t s r s t 2r ï s t s s t s s q rt s r s stér ï s tr s r r stè t é 2 r é r stér str è s str t str t t st stér t s r è s t té t st stér ré rs r t s st sé rété r q rô r t s é st s t s ré t r ss rtér s r s s2 t ét s à rt r stér s s r s q s t sé rétés s r és à 2 à t r s s t s t é rés s é s s t r s s s é s t s r ss s t r s t r t s r tt t t té s s s t st t s r t s r ts st 1 st 2 tér é t q s s r s r q rt st r t r é t 1 rété à s 1 q s t s r 2 és s t é és s s è s q st t ss t 1 rét stér t s ét 1 t st r r ét 1 t s é s t 1 q s r è 1 è 1é t q s s é s à é r s t té s s é t 2t s t é é s s s s2stè s 32 t q s r tt t tr s r r s é s t t r r é t s 2t r s P 32 s ét 1 t s t s 32 s s s r tt t ét s r s é s t t t t 1 q s s é ts é s s t 1 q s t s t é és s s r s s é s s s sé rété s s s s t st s t é é s s s s s s r ts 2 r s s
25 s t s s t r rsés s s q s è sq 1 r s s s s t é és r s r s ss r r 1 ét s t ét té é t st té à r ét r s r t s s à ét s s t 1 ét s r tt t tr s r r ét ét é 2 s ét2 r tt t r r P s s r s s s tr 2 ér s s ts t 1 q s s t és à t é ès t tr î r 2 2 é à t t s2 t ès s tr 2 ér s tr î t 2 rtr 2 ér é à t rt r r s s q s s s t à t r sté t s à t 1 té r s é t 2t s ét é 2 à s t s 2t r s P t t r ér r é t tr s t 1 q s s é ts t s t r s q r ç t s rt r t st s tr st t 1 sé à s t è s r és t s é r s tér s t è s t r s q t êtr t 1 q s t s s é t q s téré s s té s r s t t q s é r s r s s t r s é tt t é t s s 1 s st s t r s r s s 2é ï s s r t q s t r t s s 2 ï s s t q s t s t rs ré s t r é t t st é t s ts t è s t s s téré s é tés é t q s str t r r 2 é t q st rt èr t r é t ré s t r t r t r st rt èr t r r s t 2 2t s t rés t s s s s ï s é t q s s t s s s ï s r t ss t 1 s r r t t t t r s t s t ts râ 1 é s és t s q t r r s t s r r s tr s s t é s s s té t s r r s s t è s t t 1 s t t st s t é q st stré t é r s r t s t ts r ts tr s s s s ï t s é t 2t s t t r t s t r s rs r 2 téré st r t té s rés t tr s t è s sq s rs t2 s r s s s t é s s s r t q s s s r r s s ét é s t r r tt t s t r sq s s é t q s
26 s t s é s s ét t s t è s t s s téré s s s té é r ss t s s 1 r é s r s ts t è s r r és s s P P t ss t r tt r s t s st P r s r s t t q s q 1 s t à r s r s P t t r s r tt r s s s t 2 sér s r s s é s q é tt t s P ss t r tt r s r té r r é r s s téré s s r té s 1 r é s à s r 1 q s t s s s ré t rs t t rs s s s 1 r s t r s r s ré t rs à s r té P tt r r t r t r t t t s s s P s s s 1 r és à s r s s té é s t s r t r ré t r s t s ré t rs s rs s ré t rs s t t t s r és rs é t t s r t r s t ê sé r t tr s 1 t s é ét 1 r s r t s r 2 s tr î t t t s2stè t r t t s r ss è s q r str t ré s t r ré t r s r t 1 r s t 1 s r t q s 2t r rs s t t t2 t q s 1 à r 1 P P t s r t tr té é t té t t ré t r r t s r té s str ss P q té t é r s t ss r t r s t 1 t s r t q t t é èr s t r t s ré t rs s r s t s ré t rs tr s r r s sés s s s r t q r ss t r r s s r té s é s s s é q s s s s t s rq rs tér s s é P à s ré t rs s r s s r é ré s t r ré s st é t s s 1 é s t s ré t rs és s r s s r s s r t q s s s t t tr t à é t s s 1 rs é t ré s t r rô 1 t q ré t r s ré s r st s s t ét é s t r s s té é s 2é ï s t st t t r èr é s tr s ts t è s t s s é t q s téré s râ à r r r s t t 1 r ss à r s r r 1 P tt r t t st ss ré r ré s t r é 2s s s téré s
27 s t s r é t r 2t s t é t ré s t r t t s tt ré s t t st t té r r t t ér q t r t t s à s r s s té é r s s r t q s r P rt s 2 s t r tr î sé rét 2t s r t r s s s ér ts s t t és t r 1 t r s r té s tér s r s 2s r s t r s s é q s s s t és s ss t èr s q t s s t s r r t t s é s t tr s s t s réq t st 2 t t rt s s t r r t t 2 t t t s s q s r2 t κ t P s t rs tr s r t s s t s2 t ès r s s 2t s t r s α t rt s s t r s s t é t 2 q t r ér tr s 1 s rés s s r t r2 t r t sés 2 t t t t à t t t r tr s r t κ r q st q t r t r s s sé rét 2t s t r s t t t t à tr s r t t r ér β β r r é t s t très s é s s é t t é t 2 s t à t t t s r s 1 s rès sé rét 2t s r s s 2é ï s α t s s t γ ss é à rés t t t è s ss és ré r t q trô ér t s 2 2t s ér ts
28 s t s é t2 s r t t rs é t2 st s 2 2t s 2t t 1 q s t és r rés t t t è s ss és ré s s 2 2t s 2t t 1 q s t r t 2t s t s q γ t α t str t r t s s s r ér t r r r 32 t é t t s s 2 2t s t és s t r r tés t 1és s s s ré r r t t à t è ét té P r tr s s 1 q s t s r îtr s é q t t è t s t r ér r s s t r t t s 2s s s q 1 s t à r s r s s 1 rs r 1 s s t r s st é é r r ss s s 1 r s ré t rs t q t sé rét r r r 32 t 2t s r t r s γ α t 1 r ss à r s r ré t rs rt r s t 2t s r s s 2é ï s st é é r r 1 ré t rs s ré t rs s s t 2 sér s 1 sé s à s r s r s t t q s s ré t rs s té r ανβ q r ss t s s s ss é s 1 s 1 r s t s ré t rs s rs q r ss t s é r s r s ér t r s s s ré s t r s t st t 1 sé s à s s 1 rés tés r s t è s t à s t è s rr t r s rt s r tér s ré é t r r r s rés t tr s t è s q t té é é s t r s t rs r rt tr 2 2t s 2t t 1 q s t 1 r s r r rt s r t s str t r s s s ï s r t rés t t s t è s 1 2 2t s s r t ér t s s st r 2 ï s r q st s è t s ré t t r é t tt ré s t r st r t r ré s t t é é r s 2 2t s t rés t t s t è s s s t s t 1t q r t s r t r t s 2 2t s 2t t 1 q s tt t ér t r r t é t r ré t tr rt t r é t ré t t t t ss r t r t t tr s ts t è s t ré s ré é ér t s t à tér t r 2 é t q é s r t ér à rt s t è s tés q t rs rs st r s r s s é t t s t r 1 P s rs é s s s t à r tt t ér é t q s s r s s
29 s t s r t q s t s s t é s t r s s r s t q é s s tr s ss t ér t r à r r t s s t t r P t 2t r s s t t q s q t sé rét 2t s ssé t t t t r β ré s à s tr t s 2s q s P s s r r s t α t q t t t t rés t t t è s r s s t é s t r t q s t t t t s 2 2t s s s r r s t tr s sés P tr q s è s ré t s 12 è q q t t t s 2 2t s ê s 2 s r r s r t q s s r r rt à s tr s t ss s 2 ï s s s t s é t2 t r t 1 r t t s st t rs s ré t rs ss és été é tré q s s r t q s rés t s 1 sé s P r s t s q s s r t q s ér ér q s t t té rés t t s t è s ré t t ér st é t t r rés t t t è s r s s t é s 1 2 2t s s t 1t t r rés tr î é è t ér rt s 2 2t s s s t é s ré t P r t t t r s r té s r t s r ré t r s r t r ré t r s s tr t t t rés t t s t è s ss t s s st t rs t s ré t P r sé rét tr î t s 2 2t s rs t ér s s 1 r t é t s é s és t P t t s q é s s rét t s 2 2t s s s s s ï s é t q s s s t é s t r r t ér t r à s r s t s 2 2t s t és t r t s 2 2t s ré t rs rés é s s rés t tr s t è s s t 1 sé s r à s ts t s q P q t sé rét r q t s 2s q s é s β P q ré t r t é r é t ré s t r t t r s 2 2t s t s s t r s t r t t 1 s s st q é 1 r ss é s és à s r s s t é s t s t s 2 2t s t és s s s s ï s é t q s ù r t s t êtr t r s t 1 r és r s s r r s t é t 2 êtr 2tés

30 é é ér t r é t t s é s s t ér s r s r ét é t é 2 2t s t 1t t t ss r r r t t à s t è s t s q s r s P r t r tt r t t t s tr s s ré s t r é é r s2 t ès γ t t t s s t s r t ê rés t t ér t t s t è s r ttr t é r ré s t r ré t à t t tér st t t 2 t rt ré t ré s à st t r q rr é ér t q s t t t è é é ér t ré é ér t st r st t t té r str t r étr t 1 sé rt s t ss s ré é èr t râ à r ér t t ér t s ré rs rs s s s 3 s r q rès rt r r st t té r t à rt r r t t s ré rs rs é s t r t é t s r t s s s s é t é t st s ré é ér t s t é ér t r r ér t s s t r s t s é t 2t s é t t s 1 t rs r tr t r r s ss é t q tr î 2 rtr s t r s s r st ts q r ttr à r r tr r s ss t t s s tés t s q q s rs

31 é é ér t té à s ré é ér r st s t q té é 3 s r s s Pr ét é t t r s r r é 1 s ét t é à r é ré q r r r ét r té r Pr ét é tt é s r t s é r t ss Pr ét é î é t r s é st P r s sé r été tré 3 r t t s r s q rs rt é t 2t r ss é à té s é t 2t s s s r ré é ér t s rs s t s t à rt r s s ré rs rs é t q s s s ré rs rs é t q s s s 3 s r rs r èr t s ré ér rt s s ér t é t 2t s t s r s s s ré rs rs s r s t été s r é s 3 s r s s é t t s r q s s s q r ér t é t 2t s t s r s t êtr tré 1 s ré é ér t s t 1 st r s s s 1 s s s r t s s ét é s t rô r r r t r 1 t rs s s t q és s s s s ré é ér t à rt r s ré rs rs s t é ss t r t ré s t r s 1 s t t r r tr s s s 2 2t s t s t rs s é q s r

32 é é ér t r é s 1 s ré é ér t té rès t r ét é t é é é ér t é t q à rt r s é t 2t s s s s é t 2t s s t q s ts t s r t très ré 2 rs t s r é t 2t s s t és ë é t t s 3 r t r tr s ss t à rs r 2 rtr s s rés s t2 ré é ér t rés t r ér t s ér ts t2 s s ér é s t st st t é r ss s ré é ér t st ré é ré sé t sq r é t 2t s tr t s st r rt à r té rt rs t rs tr s t t t r r r t à r r t t t s é t 2t s tr t t s t ss é t q r s r r 1 s s r s r 1 st é s s ré t t t t ôt t s 2s q s s r t s é t 2t s s t q s ts s t s s s 1 ts t é q s rès és s s ré é èr à rt r s é t 2t s r st ts q s s t ç tér t s s r sé rèt t α t r r s s t r α t q t tr r s é t 2t s s 2 r t s

33 é é ér t r t s s s 1 t rs r ss r ès s r èr s r s q s t és s s r sé rèt t P 2t ttr t t r t q r r t s s t r s 1 r t t r t r t s s ét é s t t 1 r ss é és t r r s à s r s s t é s q t 1tr s t s s t r s t t s r s tr î s2 t ès ét r té s s tr s P t é r α r q t r tr 1tr r tr î t ér t 2t s t t rs t é q s é és s tr sé rétés r s s ét é s r t 2t r t t r r r t t r t α r t r t s r s t s rt q s s é t 2t s s t à és é rès q s t s rès é t t s é t 2t s s s s s 1 t rs t é q s t s q t α s rt t q s t tr t s 2 r s r èr s t s s r é s s s t s q s t é t t s t t t κ r t r r t κ s t s tr s r t t r tr s r t s s t α t κ st t é t s rès é t t t r st t à r s st t é à r s rès é t t t rs st à r s t t r s è t s r té s q é s s é r t tr s t t é t s é t 2t s t s s t é t t s s 1 r ss s è s ré s ts t r 2 s t è s ér ts rr s t à s r té s très r é s t rs tr s r t r té s sé rété s r té s r s P r s è s q t t s r r t r s é é ts
34 é é ér t ré s r κ t s tr t s è s ét s s s s t s t s 2r t r 12 s q r tt t t r é st s s t ré é ér t t s t rs t é q s t s q t α st t r ss t sq t t t è s s r s t q èt t ré é ér t s t à é t t s r2 s tés s t s s 1 è ét s st t t è s s t r s ts 2 r 2 s q t tr tr s r s t rs t t t q s r 1 t s è s q trô t 2 r 2 rès é t t s r ér t s s ét é s s s r ès è r st r s t s é t 2t s tr é s st é s s t r r és ré ts à s t r s s s ï s s ss é s 1 é t 2t s q èr t é t2 2 r st q rs r ss s r t r é t2 q s t t r t t s q èr t s r tér st q s 2 r st s 1 r t r 1 α t t2 s r ss α r s t s t s s ét é s t é s 2 r st s sé rèt t è t2 s P t tr 1tr r à r 1 té s s t s és rt t s à r t s tr é s é t 2t r s t à ré r s t r 2 trô rrêt ré r t rsq ss é t q st r st t é s êtr t é r β st sé rété r t s t à és s t t r è s s é t 2t s t 1 t rs β s r rés st ts 1 ts β rs s ét s t s ré é ér t ès q s t rs t s é t 2t s t s s s 1 ts t r ér t s β t ré t s rrêt rsq ré r t st t r é s 2 r st s r t s s ét é s t é s tr t t s é é ér t é t q à rt r s ré rs rs tr ré é ér t é t q été s r é 3 r t t s r s à rt r s s s q s t s s ré rs rs tr é t q s s s t été é r t s r r èr s 3 r t r r r été é tré s 3 s r rs q s s t s ér r é t 2t s s r s P s ré t 3 s s é r t s s ré rs rs é t q s s r tér st q s s r s à s s s s t été s r é s s s é t t s t s t s s é t t s r q s r r é r q
35 é é ér t é ét q tér t s s r s rô s s ré rs rs é t q s s s s s été ét é q ré t q rés s s rs é t t s t s s r q s s t s s rs é s r s s ré rs rs s r é s s s é t t s s st rré é sé ér té s r tr t r ss s ré é ér t r st à ét r s é s s s P s rs è s s t t t t sés r ét r r ér t s s t r tr t r ss s ré é ér t s ss t st s ré é ér t t r rt é t 2t r à t s s é t 2t s è s t sé 3 r t st tr t t r ét2 rè ss é à é t t s à t tétr r r r 4 q ré t s é t 2t s t q é t t t tr î t rt é t 2t r tr t t é r t t2 r s ré ér rt rès q q s rs s s t t t t t 2 r s s t s 3 ér rt s r t s t s r ss t rès à s s t t s 3 r t é é r s é t r ss ss é à ré t t r t s q r r t s t s é t t t t t r ér t r s s s q rs rès t str t ré t r é t t s é té ét st s è q s r ér t s s s ss 3 r t q 3 s r s é ré t s2 t ès s t 2 é ss r à s2 t ès t à sé rét s q r q t tr 2 ér s s t ét t r ét é s2 tr s ér s ê r 2 è q t str ss 12 t t é r s é t 2t r t s s s ré ér rt à s s rès é t ré tr s è s t été t sés t t 3 s r s str t t 2 t ss é à é t t s q r q é r s é t 2t r t r t s s à s s t t 12 r 2 2 r s t t
36 é é ér t r s s s q s s s t été t é s s s s r r st é tt r s s t s à ttr é s s q s t è 2é r s r r ér r 3 s r rs s s s r ss t s ré ér rt r èr t s tt 3 s r t s t s ér t st s q s s s r t r t t t t r ér t s ré rs rs t s rés t s s s 1 r r à t r tr r 2 é t q t s s s 1 r s P t rs t s éq t t r sé q s s s ss t r r ér t s s s ss s q s r t t s rés t ts t s été r és r tr s éq s s è ré é ér t é t t s s tr s è s t s s 3 s s r s β t s s β 2 t r ç tr s t t ss s r t s r s β s s s s à r t ré é ér t rés β ét t ét té s é tr t q s s s s r é s ét t s r é r ê ét é tr t ss q s s s ér t s s s é t 2t s s rés t ts t été r és r éq t q tr s è s ér ts q tr t s s é r s à r t s s st r r st s q s s s s q s t s s t é s s s 1 r s t à r s s s r é r st t t s s t t t 1 s s ù r ér t s s s 1 r s r t ss é Pr r étés t r tér s t s s s t r s s t sé q t r s r rs st r t s r t s t s t t t 2 r t r rt t 2 t é é 3 s s t é s s r é t rs é t q s s s s t s r é s s ré ér rt t s r s r t s à èr é st sés r t ré t t t r s s s 1 r t s rq rs r és t r té s t s s é q s t èr t s s s è s t 1 st s rq r s r tt t t r s s t s s r é t rs é t q s q s t étér è s 3 t t s s s è s s s s 1 r t à s s rq rs s s s é t ïét q s t t s rq rs r s s 2t ér t s t
37 é é ér t t s rq rs é t 2t r s t α t r té P s rq rs 1 r és r s s s s t é t r s é t st s q s t s ré rs rs s s r s t s é t 2t s s t 2s ré s s rq rs s s è s 1 ér t 1 s tr r q t r s s r s rs t s étér è s s q t s s ê s tés ér t rt s s rt t t s r é t rs é t 2t s t tr s s 2t s r t 1 st rq r r tt t à s st r r t s s ré rs rs s é t 2t s s s r s t s s é t ïét q s s rq rs t sés s r t r tt t ttr é 1 t2 s r s 3 s r s s rq rs é t t sés s t t rq r t ttr é s 1 r s t s s ré rs rs rq r s été r tér sé s s rq r s s r s t t s ré rs rs s r q ré é é r 3 été tré ré t q s t s s r és s rr s é t q ét t r s s ê str t r t r 1 st s ré rs rs 1 r t r t é à é s s r t tr s 1 r t s rq r s s t s r s rq rs é t 2t r s ré é t t s é t 2t r s s 1 r t q rq r é s à s 1 s r s t été s éré s s s ré rs rs t t s t 1 st st t t s 3 té s s s à s ér r rs é t2 r é t 2t r été é tré 3 s r rs s s 1 ér s ré sé s t tr t rs tés ér t s t s s t r 1 s é t q s r 3 s r ts r és r s s ê é t2 q s s s s t s r é s s ré s s s s t2 t st t été s r é s s s t rs é t q s s 1 s s ér t rs st s s r s s r t t s ér é s q s s ré rs rs ss t êtr s t s r s r s t ér s r rô st tr rsé t r t s r ré é ér t é t q st s sé s t s s rr t êtr à r t rs sq s s s s t s r tér st q s s s s t r s rt t ré 1 r ss é t té ér s t s té r t très rt t s s s
38 é é ér t ré rs rs t été r tr é s s s 2 rs ré é s q s 3 s r ts tr tés r t r è s s tr t ts t sés s s è s 1 ré é ér t à rt r s s s t r à t r à r t t rs é t q s 3 s s r é t rs é t q s s t s r é s rs s é t t s r q s t t st rr s q 1 s à r sq é é é t r é t r s s r t s ré rs rs sé s s 1 r s ré ér rt rt t é 1 st s s s 3 t s s str t r s 3 r ér t s s à rt r s s s s t s s s r rsq r ér t é t 2t r st é r ér t rs é t2 r é t 2t r q q s s rr t st t r t r t r ré é ér t t t r tr t à ré é ér t rs é t t s r q s 3 r st à é tr r t r rô s r t t rs r st à ét r r é s s ré é ér t à rt r s ré rs rs s s s s t t t r t t t s q s sé r s s ét é s s 3 s r t r s r tt t s t ts tr s 1 t2 s r s t s r t s ré è ér t t r s s s é t 2t s s é s s t t r ér t s s s à rt r s ré rs rs q s t s s 1 r s t r très s s s 1 t t r r t t s é t 2t s térés r t r é r t t s s té é t s s ét é s r s rt q s s é t 2t s α t t rô rt t s t t r ér t r rsq 1 ét s q s s t α st é 1 1 ré é ér t r s s ré rs rs st é 3 s s r s t t s r ré t r α r ér t s ré rs rs st é q t rô r α t ê 3 s t ts t 2 t 1 α α t r t s ré rs rs é t q s st s èt t é tèr ré é ér t à rt r s s s s ç s s t r q s ré é ér t à rt r s é t 2t s s rés t ts tr t t t s t rs r ts s t ré é ér t à rt r s ré rs rs s ts t é q s α P t s t s
39 é é ér t t rs r ér t s s s s é t 2t s r s t rs s t érés rs r tr r s ét r té s s q t r t très ré t t r sé rété r s s ét é s é t rô s r t t ér t s s s ss été q é s ré é ér t tt 2t sé rété r r 1 t2 s r s t s r s s é t 2t s t s s s t r t r ér t s é t 2t s t s s s tt ré s s r t é é r t t r s t sé rét 2t s r s r s s s t s t été s r é s s t r ér t s é t 2t s t s s s ré t q s êtr é t s t rs q és t r r t s é s s r tt t t r s é q t r ér t s s s ré s é t 2t r s t s r t ét s rt s t rs rr t r s é q t s 1 t2 s r s q t2 r rr t ré r ér t à ê st s tr 2t s s t t t s r r ré s ér t 3 s é t 2t s t s s s 2 t 1 β β t r ér γ t β r α r t s s st sé rété r s r s t s s t q st r ér t s s s s s t r s é t 2t s r 1 r ss s ré t r s s r s st t rs ré é ér t st 1 r é à s s s s s t s s r s st ss té 3 s s s rr t q s t s s é t t s r q s ét s r é s 2 t 1 β st tr s 1 r é à s r s 2 2t s q st r t s é q t ré s ré é ér t à rt r s s r ér t s s s st ss é à t t s2 t ès β rs q tt s2 t ès s ré é ér t rès é t t s t è β ré t r ér t s s s 3 s r s s rés t ts tr t t s é q β s r s s s q s ré t r s t ss 1 r é r s é t 2t s t q t r s r ér t s 1 t2 s r s r ré t r β st é t t é r 2 t 1 1 ts 1 r ss t s t 2 r t r r t r 1 r ss 2 2 2t s r q st r ér t s 2 2t s t ré s é t q rsq β t s s t ré t r t ré s r ér t s s s rs q rés
40 é é ér t t ré s r ér t s é t 2t s st r rt β q rr t trô r s 1 ré s s ré é ér t s tt 2 t ès s êtr r é sq ré t t s éq t tré q β t é t r rô s t s r ér t s é t 2t s r t t r t t r t t s2 t ès t r ér γ γ st 2t sé rété r s 2 2t s t t 1 r ss st t rs r ér t s s s t é rs é t t s tr rt t è ré t t s s s 3 s r s rs q st ré é ér t é t q rès é t t s tr t t r ér γ r ér t s é t 2t s t r α t P rs q st s s t s r s s s s s s 1 ê s st s s rés t ts s t rt γ t s 2 2t s s trô ré é ér t r β tr s r r t t r β té r r ér t s é t 2t s rs q s s s s t s s s s à t t r tt rés st s é à 1 r ss s é é r t r t t r s s t β s s s ré rs rs r t s β r s r s t s 2 r st s r t 1 s s t r ér t s r t t r s s s sé rèt t r s s 2t s β γ t s 1 r t s ré t rs s ts t r s tr t à r r ér t r ér t t r r t s é t q s str r t r t tr t t ér ér rs tr rô rt t s r t s s q 1 r t ré t r ré t r s 2t s t t r r r st t r t r s r s t s s s st ér t s é t st s é t 2t s rs é t rr t r ê rô s t st t ér t s s s rs é t2 é t 2t r
41 é é ér t r é t t ér ts t rs s à ré é ér t à rt r é t 2t s r s q 1 ré t ér t s 1 ré t s r é rès t s 2t s q trô t ré é ér t é t q s t sé rétés r s s té é rés ts s s r s s q s t r r té s s r èr s t t tr s s s s ét é s q sé rèt t t t s 2t s s ét r té s s t s r té s tr t t s s ét é s t é ôt tr rr t ré é r 1 s s s s été tré r tr s s s ré rs rs s t ts r ts t r t ss tr s s t s s s rès r t r s s r t s q s t r t r ér t é ét r s rés ts s r s r t r ér t s s s rès str t s s r s tr rt s s q sé rèt t r s s 2t s α β q é s s r r t t s t r s r t s t t t s s ét é s t s s 2 ï s 2 2t s rô t s r ér t s s ré rs rs é t q s été tré r s ts t rs é s t t r s rt s 2 2t s st ré é é r s ts s é q s s r s s s 2t s sé rété s r s s γ β t r t tt ré é ér t 3 s s r s 2 ï s
42 tr s é t t s r q s st r ét 1 t 1 sé à s t è s à s é s t 1 q s q ét s t é s s é s t s ré é r t 1 q s t tr î r rt s é t 2t s t tr s s r 2 t s s q t s s té é r r t és s r s r t s s 1 q é t r ss s r é é t s s téré s t ré r t q t s rs t2 s r s r ss s ré r t s és s s t s rs ét s r s s téré s é tt t s s 1 t t s r s rés ts t s 2 2t s t t t r é t2 és s s rés t s t é s s t à r t r r r t t 2t s r ts s é t s s téré s r 2t s t sé rèt t s 2t s t t r ér t s é t 2t s α t tr s r t s s ét é s 2 r st s β r ér t s é t 2t s t r és r t r té s tr r s 2 r st s t é t t r r t t ré rs rs s t é s r r r r s s s ï s r tt t tr s t r rs st és s s à s ts t è s é s t 1 q s 1 s r s tr î é t é t t s r q s q rés t t 1 st r ss s és s t ré t t r t r q ré s à rt s s t t t t s s r é q s s ét é s t r st s rt 1 s r é t2 2 r st q t tr t t ré r t t ss r rs st t r s t 1 ès tr 1tr r r ss s r é q s s s t ré s ré é ér t q s st r t s ré rs rs s s s t t s ù té r ér t s é t 2t s st é

43 r é é ér é t s s r q s s t s r s t t s r ét q t 1 q s s é t 2t s s t s r s s s s é t t s r q s s t 1 ss t r q r st r èr s s t s é s à t r ss s s r t s s s tr s 2s é és s s t r r é s s s tt2 r s s s t t s s réq t t tr 2 ér s é t q s s s tr s 2s é és st és té ss é à rés st à s t s s t s é t q s é s à t s t êtr r é s à t r à s t s t r r é s s s tt2 r s s s ré s q q s t st é t st t t st t s s 2s t 1 t st st é tr t
44 sté t s t é ér à s rs s ét q s s rs s ès s s ès s r s t s t ts ès s r s ét q s t s t q s st t s sq 1 st s ét é st s t s s ét t tr s s éré s rés r é s à t s s s s t r r ss t ét s à t s s â s t t t t t ê s ts q t t t t r r t r té s s s ét s t r rté ré s r t é ér é tr q s 2s t 1 t s ts ès s t t ré és té t èt t2 s t 1 q rt t t r s t t é q s t rés t t s és s s s t t êtr st é s st q t s s é t q s é s à t r ss s s r tt2 r s s s r r t t t s tr tér t s t s t s s s é t 2t s sté t s é t q à sté t é t t ù ré t t r s t 1 tér t s é t 2t r s q s rs t r s é t q s rs st t t s q r rr s st t s éré rré rs 3 sté t s s s r s st s éré é rés r s st ré t à s r r ss t t s é s s ér s sté t s sté t s st t s ss t t s tr 2 ér s à tér r s r s q t t t t s s très s q t tés s sté t s é t q s tr 2 ér s s t s s s 2t s q s s s é t 2t s st st sé rsq t q é ss à s ss r q t sté t s t êtr r r s t r ss é à s t à és té èt t s rt s ét ts tr t s r r r r r t s β 12 t t r r é ét q t 1 q s sté t s r r 2 s q t t st s ér r à 2 q st é é ér ér rs sté t s r r t 2 r s rs s q s t 2 st s r té ér ér r é t sté t s r r rs t2 r r été s r é r tt t s s r q s s t

45 sté t s s r r r r s r ss s s s rs st é ér t sté t s r r q st s r é r s ss é à sté t s r r r s sté t s q s r t é t 12 é s 1 str t sté t s r r à t r r à r t rès s t tr é t r ô t P r ss à tt tr t r r r r st t s 1 t s tr 2 ér s 1 é t r s st és s s s é t 2t r s t r r s rs s r s rt t r r s t 1 ss rr t r r r ss s t t rr é s r s s s t à 2s t ss 1 s és té t s rés st t t s2 t ès s tr 2 ér s s ès à rt r s r s s2 t ét sés t 1 rt s s s s t 12 t s s r s sté t s rés t t t q t té s r s s s à éséq r tr s tré s s r s s rt s r t t ès t r é t r β 12 t t r sé rét s s r t s t t s s t s st s r s tr r q 3 s t ts tt ts r s tr 2 ér s é t q s ér t s r s r s r t ss 1 s2 t ès t t t s rés t ts tr st t 1 t s 3 s s té s ù s2 t ès tr r s à r t s tr 2 ér s é t q s r é s r t sté t s s s st rés st à s tr s é s s és à s rés st s t ss à r sté t s t 12 t t r r s2 t ès α q t PP γ t t r ès
46 sté t s rs sté t s s s r s s t r t s è s ré t s 12 è r r t 12 s s t str ss 12 t q r st té r st s s r s à tér r s s s s é t 2t s q t êtr s éré é s r t t s s té s é t 2t s st rés r é s s s t r sés r t r t r P t r té ré t s és s é t q s s t t q t té s r s s 2t s t s s r s tr î t é r s t s rt s é t 2t s rs é r s s tr 2 ér s és s ré t t r rt q té 1 s r s à très î ét sés t s és s t ss é t q r t rés é sté t s r s é t 2t s s ér s à s tt t s t 1 q s t t r té ré é ér t st t r r sq r é t é t t s r q s r t t s r t tr t à é t rs sté t é t t t rs trô t ès é t q st t s2 t ès s r s s st é é r s t rs tr s r t s P st r r t r2 t r t t é r s P r 2 r t r s s t r t t é r s t PP γ r 12s r r t r t t r t r γ è P s t rs s t s t t rs s è s ès t2 r 12 s tt2 s2 t s t st r 2 s t r s ét s r s ès s é t s P tré ré t s tr 2 ér s é t q s t t 1 r ss s 32 s ès t è P 3 s r s tr î t 1 r ss s t s s è s é q s s s 1 r ss PP γ st té s s 1 rs sté t s q tr ss à s2 t ès s s é t q s st t s 32 s ès s ét s 3 q t rô rt t t t ès s st t sté t s é t q s s è s 1 èt s rés st t t és té s ét s t s t s s t s st s 3 s t tr r q s2 t ès s r s t r t q st t s q t té tr 2 ér s s2 t ét sés r s s s rés st
47 sté t s t rs trô t 12 t s s r s t sé rét s r t t té 12 t s s r s t é t êtr r s s r t sté t s é t q s t r q s t r s t tr s str t r s r s q 12 t s s r s t r s sté t s 12 t s s r s st rré é à q t té s t q s r s érés r t ss 1 t st t é s tr s r s s t r s t s r 12s s ù s t β 12 t t rét s q ù s t ω 12 t tér t s t r s r s tr î t β 12 t t t t ω 12 t r s q é èr s s r s r 12 q s q s t s s PP α PP α s rt ét t r s s s r s r ît t ré à 1 s r s s t tr s r t s è s q és s 12 t s s r s P r t t 2 tr s r s q st 32 t t β 12 t t r t r t tré s s r s s t r s s r s é t s r PP α é t sté t s t sté t é t t s r q s 1 r 1 s s rs è s s r q s PP α rô r s t ès sté t s t s é ér t s trô t 12 t s s r s t ré s t r t 1 té s tr 2 ér s s t r rés s s s é t 2t ss és à s s s t à r té r r té tr s rt r s P r s tr s r r t s s t s t 1 rtés s tér t s s2 t ès t sé rét P t ss êtr é s s t à t té 1 rt s tr 2 ér s à r t sté t s é t q s s s sté t s sté t s é t q t êtr r t 1 q ét q r s sté t s r t 1 q st s t r q t 1 ss ét sé r s é t 2t s st à r r t q tèr t s t r s t t é t s s r s r β 12 t t ss r t t r é té t st t t tr t P s s rt q t s s r r t α t ès tr s é s t 1 q s ét ss s2 t ès ét t é s2 ét s s t t r t s s s ts tr î t t r r t
48 sté t s tr 2 ér s s s t t r sé rét s s r é t sté t s é t q st r ét q st s s t é à és té èt t à rés st à s s ré s s str ts é r ét q s s s s s t t ss 1 rés st à s ér ér q t à t té s s r s s s r s t s 2t s q t t s s q t à t r 2 é r s s rés st t ér t s s s s t à t t é ès t t s2 t ès 2 è 2 r 2 é st s2 t ès s q tr î 2 r s é t tr t t s rés st 2 r s é st ès tr é t q β 12 t s s r s t t r èr t s t r 1 rt t s é t 2t s r s s t ss 1 rés st à s t 2s t q t té s r s érés s s q t à t t q t té s r s s sq r t t 2 st s ré é r éséq r tr t t t t t s2 t ès s s r s rt t t té à s 12 r t à s 1 rt r tr rt t t à r t sté t s r é ré t t s é s s s t à sté t s é t q

49 sté t s à sté t é t t s été r t ét s st s rés st q s t tr 2 ér s s s st rs sté t s rt r r t t s s s 1 ts r s t rr t êtr r s s s rés st é t q rés é s 1 é s s q s t t s t tr t à sté t s t à r r ss rs sté t é t t sté t s à sté t é t t sté t s st s éré t é t ré rs 3 r r ss rs sté t é t t s à s s s s 1 ét s s t s t rs r t sé s s ét s r é r s é s s q r tt t tr s t sté t s é t q rs sté t é t t s t sé s r ss t s r s è s 1 q r r s t s s s r tér st q s t s ét s 3 s t s r 1 st s t q s sté t é t t r q s r tér s r sté t s r r s tér t s é t 2t r s s t q té é r s r s t t q s t tr t s t r s s r s r2 s s 2t s q s ts 2t sq tt t êtr s r és r r st q ét t sté t é t t 1 rès s t tr é t r ô t P r ss à
50 sté t s à sté t é t t è s è r sé r 2 s é t sté t é t t st séq 1 é è ts s ss s t t r r st t tr 2 ér s s s é t 2t s sté t s q s r s à s s é t tér t s é t 2t s s s st é ér t 1 s t à str ss 12 t à s 2t s t r s rés t t t t s r s rés ts t st r s st r s 1 ét s sq sté t s rt t t é r s tér t s t r s str ss 12 t rt s é t 2t s t t t ré t t r s sté t s s t t q s t s r s q t tés rt r s r s r s stér és ss êtr à r str ss 12 t t t q s t à r r r ss sté t s rs sté t é t t t rs q és s s s ér ts é s s ss t s2 r r ét s s ss s t été r sés r 1 q r tr s t sté t s à sté t é t t s t rs q s t ét r ts s t t t ér té s é t 2t s sté t s q s r 12 t q t str ss 12 t tér t ét s s s 2s t t t r t s rés st tt r èr s t s séq s ét q s é s à s rés st t 2 r s é s ss r t s r t ss 1 ré t ét t r t r t r t r r t q r 1 t rs s t q és à s s t s s t êtr r és 1 r s té r s s t rs ré t t t str ss 12 t t s t rs é t 1 r ss 2t s r t r s t ê s s t rs t t r é t r rr t êtr r t r t s s é t 2t s 2 t sté t s rt t t ér t sé t tr 2 ér s t s r s t 1 q s s r ss s s tér s t st t r t ét t èr P q t r t α r s s r t t t é t q t t r tér s s t st rê t t t r é té s r s
51 sté t s à sté t é t t t st s t été é r t s s réq t 3 s t ts tt ts q s t é ér s és s s r s t é t êtr s é s r é t 1 q 1é t q t 1 t s rt s 2t s α s é ér t str ss 12 t q t r t s è s ré t s 12 è ô str ss 12 t s s s str ss 12 t s t r tr és èr st t s s s tt ts sté t é t t s rq rs str ss t été r tr é s s s s t ts sté t s é t q s t èr r s rt t s s s s é t q s s tt ts str ss 12 t st é s tr s r r ss sté t s à sté t é t t sq r ér t s é t 2t s q ré t té r t s s rt s r s r t r s 1 s s ré rs rs ss é s à r s s é s r 12 t s s s t s s sté t s q s t s é s 12 è r s s s s r 12 s t s r 1 2 r 12 s s s t é t r r és s s t s è s ré t s 12 è r r t 12 s s β 12 t s s r s st é s r tt t r r é r t t r q t té s r s s s é t 2t s s é t r r s s t s é s 2 t ré rt q r s t s s ss t t 1 s t ss tr î r r 12 t s s t rés r s t s é s ré s t r r t s r s és s à tr s s s s ts t 1 q s r 12 t s s s r s r s r s s é 2 s r és rs tt ré t tèr t s2 t ès s é t s t s r té s t t q t té t t q st t 12 t str ss 12 t é t t s r s t t à r tr s r t s s ét é s 2 r st s r

52 sté t s à sté t é t t r é ré t t rô s s è s ré t s 12 è s é t sté t s s à sté t é t t t rô r t β 12 t t r t s é t été s té sq st t β 12 t r t PP α s è sté t s 3 s r s tr î s r t ré s r ss s é t q s s é t t str ss 12 t s q ré t t t r s q q s s s r r q s st t st s s s s str ts r r 12 t s r ss s r tt t ré t é t sté t é t t r r r r t r 12 t s s 1 ès é t t r sté t s st rt t ss é à t é t q r q à t t κ κ t à é é t 1 r ss 2t s t r s α t β κ κ t êtr t é s s é t 2t s r t t r s s r s r s t κ r r t r 2 t s r r étés t 12 t s tr î é r t s t
53 sté t é t t à r s t t r s s è sté t é t t P r tr t α t t r κ ré t t t κ é t sté t é t t s ê è s rés t ts tr t q str ss 12 t t t t κ s t s t t rs s2 t ès α t t s è 3 s t ts tt ts sté t é t t q t té α st té s t sér t s 1 tt 2t s s t rré és t st q s s rô r t r α t s rés st rs q s t é r s s té à s t s r ètr s st q s és té tr ss à t ss é à sté t é t t r r t t r s s t ss 1 t r t α t s t rés st à s t 2s s t ss 1 s ts t t t 1 sér q s r s r s t 2t s t r s r t s q s t s t rs 12 t é t q s s r s P r è t t 1 sér q t tt 2t tré s s r s s s é t 2t s s s à ès t st β 12 t r t t P s t PP α s t 1 s t s r és 3 s t ts ès s tt ts t rt r à t t sté t s t t t t r t rs s ts α é r q t α s2 t ès t s ts t s t t ss 1 r rt tr t tr α t t st é é 3 s s r s ès s é t s t t st ét r t s sté t ès é t q 3 s t ts ès s tr t s r r étés t r s t s t t 1 sér q t st é é t st r r ss sté t s sté t é t t sté t é t t à r s r s é t q st t 1 ss tr 1tr r r tr é à st é q s t t t s s s r q s rr s st t st s r r s é t q t tr î r s s é t q t 2 rt s rt s é ès s t rs ét q s ét r t r r ss s rs r s s rr s s t r s r s s st é t r é ôt tr 1tr r ér s s ï tr r t ér rt st é s é ôts st t t s s t r 1 q r t s ts tr s s tr r s t s s s rt 1 t tr s

54 sté t é t t à r s s s rt 1 é t r s st é é r 2s st q s r q é st s s r s r s ér s s ï ér rt s s s t r r s rt t ér rt r r s s t r s rt t ér rt r 1 s t rr s r s r s ré é ér t t r r t t r é t q r r s st q s r 1 r t é t 12 é s s r tr t st r s é r ss t s rt t é ôt è s s r t s t sté t s st é à r t r t r r s tr t st rr s r t t r st é s r s tr r s r t r t s s ré é ér t t s t t r 2 t r tr s s tr r s t s s s rt s st r t sté t s s r rès s t tr é t r ô t P r ss à tt tr t r r r r r s 1 t s r 1 rt s é t 2t s ré st s ré é ér t q s r t t r r r ss rs rr s r t s ré é ér t t r t s téré à r s s é t q st s ré é ér t t r t rs q s é t 2t s t té r ér t téré r s t s r ér t s s ré rs rs 1 s é t 2t s é ét q t térés r s r 1 r s s r 12 s q s s 2 rs ré é ér t rr t êtr à r r s é t r s sté t é t t q t à t é ér é rs r s q q s é s s s s t sq st rr s r s s é t é t r t r r t s s s s rr t q s s t à s r ér s

55 sté t é t t à r s é s s r ès r s é t q st à r ss s ré r t 1 éré r tér sé r t 1 ss s tr 1tr r téré ré s à r ss r q rés t éséq r tr s2 t ès é ôt t é r t s s st t ts tr î r t r r ss tr 1tr r r s ssé r s r s è s r r t s r t t r é t q rs r s é t à és r q s é t 2t s ésés tr t t s s s t s tr t s r 2 é t q t s s r s t t r â t s é t rs r é q s s s ét é s s tr s r t 2 r st s q r èr t t r s t r s q t tés r té s tr 1tr r t é s s s ï s st s stré t s tr t s s s ét é s t é s tr î t t t rés st 1 s s s s s ï s é t q s rès t r t r r rs s tt t s é t q s ës és t é ré t t r s ré é ér t ss é à é ôt éré tr 1tr r r ss s r t tr s t és r st r t r t t r é t q r r rs és r q t
56 sté t é t t à r s rs st t t t r ér t s 2 r st s t tr î éséq r tr s2 t ès 1 ss s ts tr s t s é s s é r t t ss t à r s s t r s é t r ss s é ét rs é t t s r s r q s s s s r s r s r ît s s 3 s ér rt s rs q s é t t q é t s 3 tr r t ér s s ï rsq r s r r ss ré é t s r s è r t s ts tr s s s rt s t tr r s r 1 s t s tr 1tr r s t q t t t t q t t t t r s q q s t ét s s s r s é q t té t t tr 1tr r st t é r s 1 st st t é ss t t è s t2 t r t é st t r té 2 s t tr 1tr r rés t à s t t s2 t ès s s s ts t t r é r t à t t té s P r t t rs t rs s é q s s P t ss t r t r t s s s s ts tr s s t s rs r ès s t r t r t s2 t ét sés r s s ét é s t é s 2 r st s s s ét é s t é s r t s r s s t s ré r t sé rèt t r s q t tés tr 1tr r t ré t é r t tr tr s s t rô s é ôt tr rt r s r st s rt 1 q r èr t t r s t ts ss 1 rt 1 rs t s r s s r st s t s s ét é s èr t r s rq rs s é q s t s r ré s à t s s r 2t s s 2 r st s ér és ss s rr t é t r rô s r s é t q s s r t r r tés rs r r ss r s é t q r r r t t été s r é rs r s t r tr t t tr s t s é t é s é t 2t s s r s s ré rs rs rs é t2 és 2 t 1 tr s t é t é és 2 t s rr t ss êtr à r 2 r st s s tt r st très tr rsé ér ts t2 s ts t 1 q s tr î t és s ré t t r r r r t t s s r s s 2 2t s r t r st tr s r t s s ét é s 2 r st s r é q s q sé rèt t 1 ss s é s t r s t tr t t r t
57 sté t é t t à r s és rs st s s ét é s t é s t s 2 r st s rt 1 s t s2 t ét s t r s q t tés r té s tr 1tr r t s t à r s é r t tr st é r t t s P t s s é t 2t s st r ès t s s r é q s st t r r 1 t rs sé rétés r s é t 2t s t s s t r s rés t s ré s à 1 s t à s ts t 1 q s t è s r P r s t rs s r 1 r s α β sé rétés r s é t 2t s t s s r t γ sé rétés r s 2 2t s t s s r s t s é t rs tr s r t 2 r st q s s ét é s q s t s s 2 r st s q q s t r r sé rèt t s 2t s P q tr r r t t s s t r s t β q st èr t r t r r sé rét r té s tr ét r té s s t rs t rs és s rrêt r s ré r ss tt s é ss t t s s s ét é s t é s t ré é ér t s é t 2t s s è s s t é r és r t t t té s P s t à t s P r

58 sté t é t t à r s r s s r 1 é s s r s q és s s s r q s t ss t à r s té rès t r t r r
59 sté t é t t à r s t s t s r s tr s é q s r s tr r rô r 1 t rs s r ès s è s ré t t s t é r s s é t 2t s r 1 1 t s t ét és t ré s r é q 1 ré t ré s t r P β γ ét r t é t t s té ré s r é q P t γ st t rs q s é s q é s s s2 t ès s è s ré t s 12 è P 12 s ré t à s t t é ôt tr 1tr r s t rs r ss r é q s β t s s st s s t s r 1 1 tr q q t t r s t t t s t r é r q s t r r s t s t s s t r r s t t t t r s t t t s rô s é t r s s é s q é s r t t s s2 t ès t r tr P P t β t rô ét r t s r ès s é s s r ès 1 ré é ér t t t r r s t r t s tr s rs t2 s r s s r t s 1 s tr 1 t à és s é t 2t s és é tt t s s è s ré t s 12 è s é t rs r é q s t s t t t s s té rés ts t r r t t s r t s tt ré t t r tr î t t s s ét é s 2 r st s q s2 t ét s t s è s t s r té s r tr sé rèt t s s t r s 1 r t s é s és t t t t s 2 2t s s r tr 1tr r èr s t rs r ss q ét t ss és 1 r té s tr s é s s tr t t st t ré r q s s té t s s r é q s rsq s s és s s r ît r s ré r ss r rrêt r ér t t é ér t t s s 2 r st s r ss s s é r t 1 ès tr 1tr r r t s P t t à ré r ss r s t à r st r t r t t r é t q ô s s s r s q s é ré s à s tt t s é t q s r r é r s sté t é t t st ss é à ré t t r ér rt tt ré t st r tér sé r r ér t s 2 t s r tér st q s s s s é r t s 3 s r rs q s t s s ré rs rs é t é s é t q s t s té ré t t r st rr é à t s té r s é t q s rés t ts s èr t q s s ré rs rs é t q s rr t êtr r é q s
60 sté t é t t à r s s t st t s 2 r st s r sé rét 2t s r é q s s t t t tr s t é t é és 2 t s s é s s r é q s r ttr t 1 q r r t r s ér rt ré s à tér t é t 2t r s r s t s rt tr tt ré t t r s r t séq ré s ré é ér t à rt r s ré rs rs q r èr t s ré ér rt t r t s r r étés r é q s s tr 1 ré ts ré sés 3 s r s t tré q t tr 1tr r ré é r t s s s 1 t t s q s s ré rs rs ss t tr r à r s rt r sé rét é s r é q s r t t r s t r t s s 2 r st s rt 1 è s 1 é t t s r q s ér ts è s 1 t été é és r ét r sté t s t s r r ss rs sté t é t t s r s t s è s r r t t t s s r tér st q s sté t s é à és té 3 sté t s t s t é t 3 s s r s é t s r ré t r r r ss s st ss é à s rés st 3 s r r ss s rs sté t é t t r s tt é t t êtr t s s 1 s t 1 sés à s str ss t 1 q t P s è s sés s r é r tt t t r sté t é t t ù sté t s rés t é t tr î t t r r t s tr 2 ér s s s t r sé rét r s é t 2t s s è é s r ss s t t t êtr ss é à t s2 t ès è à rt r ét tt t st t s t s r t ét t t ré é t ét t s t q t ét 2 s tr s ér s r t è s t r t s t t ét ét ré é t ét s é té ét s è s r tt t t r sté t é t t s s s rés st s r é s sté t é t t é à és té 3 s s t t très t s r ét r sté t é t t t r r ss r s P r s è s ré st s t s ts t 1 q s ét t tr r t è r t t r r s ss é à ré s ré é ér t s tr à rt r s s q st s r é 3 s t ts tt t P r t r r s é t q è s t sé st t tr ér t é
61 sté t é t t à r s ré été 3 r t s r s st ét sé r s é t 2t s tr r s é èr s r 1 r s s t à r 12 t s s à r t s t é r s s é t 2t s t ré t t r t s é é r t t s s r r s t s r s t t r s é t q s t à s s 3 s r s t é ét q tr è r s s t sé t é r t st str t t ét r t tr ér t é é s ss s 1 s r r s è s t r t s t t t r s é t q s t st t à s s 3 s r s
62 tr r té s r té s r t rr st s st r té sé rété t t t é r t è t s s r st s rrêt r ss rs t r s sér s2 t ès t r té s t t rsq r ér t r st é s t r q sér s t r t t t s s t r s r s rs 1 r ss s rsq 2 r r r s 1 r ss r té s été ét té s r 1 r s t s r t 1 r é s r s r té s rt t à r r té t t té é ss t γ r 12 t rés s t q s é t t s rs r té s t s st ré t rs à t2r s s sé tr s r s 1 r t 2 tr t r t 1 r ss s s é t r s s t été és 3 s r s s 3 r ê r 3 s r s t 3 sés s r r s 3 s r s t 3 str t r r té été t à rt r s séq s é t q s st r té sé rété s é r 3 t 3 s r s st st t é s r 1 s ré t r r s t q s tr t t t s ré étés t2 r s 2sté s t r à 1tré té t r sé t s t2 s à r té s t q st r r q t r ss t s ré t rs r r s 2 t t té t2r s s r
63 tr t r t 1 r ss s tt str t r s st très s r à r té r tt t s é r st q st t r r té tr t s 1 séq s r té q s s t t q s à s tr r t à r té s t t s s t r r s sér s r té s s t r r r té s r s t tr t s s t2 st é ss r à s t té t t r tr t r r té s s 1 è s rt s tt s q t s s t s t s t r t s tr s rs q t s s t s 2 s2 t t t s s t s t r t s s s ré t rs rès rr r P r êtr q t t s s t r té t s r t st tr t s rés s t q s q t êtr γ r 12 és r r 12 s rés t s rét s q tt ré t st ss q rés t t tr î t r t r té r t s s t2 ss s r ré t r s t s t2 r tr és s s r té s très rs s t rs r ss r té s és tr tr s r tt t s t r t s tr s r té s t t rés t r t s s s r és é t t s r té s t s t êtr q é s s s t r t s r s sq r r r ît s ré t rs r r s t2 tr s r té s tr s r r s r té s és t s s t γ r 12 é t t r r s s s

64 é t rs s rés ts à s r s r s t t q s à s r s q tt s t é s 1 r ss s t é é rt s s r st s s r s rrêt r ss été s r é s r 1 r s r ss s r st t s ér ts t2 s r s r st s s t é s s 2é ï s s s r s ss s sé rét s t s t 1 sér q s t tés s rt s rs t rs és s s r s s r 1 r ss s st 2 st sé rété r s r s rés ts rsq s s ét é s s tr s r t 2 r st s 1 r ss s st t s s s é t rs s s st r s ré t rs 2r 1 r q st t t s s ré t rs à t té t2r s s r tér sé r t s r é s à t té t2r s s P 2t s q t ré t r 2t s q t t s s t2 t r t rés t t s s t s 1 tr és s rt s é s és r r rés t t s ré t rs t s t r té s t s s 1tr r st sé 1 s s s 1 s t 1 s r t t2 tr r t t str t r t2 q ré t r t2r s s rès
65 é t rs s t 2r 1 r 2 2 s s è s s é q rt s r s t s r s s 2 t s r s t r s r s 2r r t 2 t 2 r 2r r t r t r s ré t rs rès r s ré t rs t été é rts t és é t s é s r ér t s éq s s ér t s s è s q t à t r s t s ré t rs s s t és és t t t r s s 1 2r t r rs è s t str t r é q r 1 s r r t 1 s r 1 t 2r t s tr s r ts à s s s t s é ss t sés s r té s t s str t r s s r s séq t s és r s t t t t té à s r rt 2t s q s r té s t rès t r t s s é r s r t à r 1 t 2r t à r r s ré t rs ssè t s r té str t r s ré t rs ss t s 2t s t r s t r ér s t2 s s t s é q s s r és t s t êtr s r rs r s s é t ê t 1 r r s rs ré t rs t à ér rt s tr s s ré t rs à t2r s s r 1 r ss t rès ss t st t 3 t s ré t rs s t 1 r és s r 1 t2 s r s s s r é t ïét q s s s é t é s s s t é s r st s 2r st s t 1 r é s s2stè r 1 s ss s s t st s s r s r s s s r s rét t s rs é s é t ïét q s t s 2t s r s t s q tt s 1 st r t 1 r é s r s s 1 é és s t r t s q s s 2t s r s s q tt s s s t é s r s

66 é t rs s s s sq tt q s s r s t s t st s s 1 é és ré t r r s t ét tés s s r s r st t s t st s s s rét s r s t s s s r é t ïét q s s 2t s r s s s r t q s s s s s s é r2 2t s t s q tt s s ré t rs t s r té s t s t s s ê s r s 1 r ss s s t ss s s èr s ù s t s t tés q s ér t s r r ré t t s str t r s s r té s t s s ré t rs s q s t r t s s ré t rs é r ts s s r s rès 3 r s st r s ré t rs 1 2r t r s st t s ss t r s t s t rs q r té s 1 1

67 é t rs s ré t rs 2r t r s té q s 2 s é à r t t ré t r 1 r r té tr t s s sér 3 à st s q r té q t tt r rs ré t t r ë 1 t s r té s r èr s ré t rs t t r t s ér s t q r q t s r2 t s rés s t2r s s t s rés s t2r s t r ré t r r s tr s rés s s t s r és s s ré t rs 1 t 2r s s t s s r2 és s t à 1 t P r ré t r 1 t s r2 t t r r t s r s rés s t2r s q s t r q t s s t s r r s é s s s t ssé t ér s t t s r r tr 1 èr s t q s ér s t ré t rs ér ts étér ér s t q s ré t rs s t 1 r és s r r ê s r r 1 s s s r ér s t t t t s ré t rs t ê s r r s r t t s ré t rs s r s t r t s tr s s 1tr r s 1 èr s été é r t s s s s r 1 r t 1 s t r t s t r r s r q t ré t s s s r rés t t ér t s ss tés ér s t s ré t rs r ér s t é t ér s t q é t ér s t étér q tr 1 ré t rs ér s t étér q tr ré t r t ré t r tr r 1 α s tr s r tr s s 1tr r s 1 ré t rs
68 é t rs s s ré t rs t ss êtr s t t r t s tr s s s s t ré t r 1 t s ér s r î α ré t r à r s s 1 t s r s ré t r tr î s r2 t ré t r t 1 rès s ss s t s s r s ré t rs à t2r s s st ss s s r q s ér ts èr s r t s s étér ér q s r ss t r s rô s ér ts r s t s r ré t r 1 s t2r s s s r2 é s r t t ré t r 1 rés s t s s t s r s r s r té s ssé t P r s r té s P s P r ît s rés s t r tt s t t t κ t 2 s2 t s s st q é s s é s s t t t q s s s γ q r ît s rés s t t 1 t r té r t s r rés r t t t q é s s ts t é q s s s r s r st s r q s s s ér s s P r s tr s rt r s ré t r 1 t é s r t q 2 r t t r s r té s s r ss r 2t s t r ré t t r t s t s t s t q t s r r s t té s t s t q r t é r t ré t r 1 r r té s rès s t t s t s ré t rs s s t t r t t ré t r 1 s t r t s rs s r tt t t r s ts q s ér ts t t é t 1 q s rô s rs é r ts s s 1 rt r t t s t rs tr s r t κ

69 é t rs s r Pr té s t r ss t ré t r 1 rès s s r2 t 3 r s t s r s ré t rs r t 2r r ré t r 1 t t ré t r r t r r t t s r s t2r s s s r2 é s P s r P γ t t t s s P s t P s t P r t t P t r t s 2t s s r s t t q s és 1 r s r t r é r s t s ré t rs s s t r ré ss ré r s t 2t sq tt r t r é r r q st ré t r s t t s r té s t st t t t ss é ré t r r st s r2 é ré s à t t r t t r t s 1 r té s rt à ré r s t 2t sq tt t t s s r s rs 2t s

70 t s s s t s r Pr té s t t r t été é r t r t 2r t s t é s 3 tr s t s ré s à t t 2r été ét é t r 1 t r s r2 t 2r t s s P s t s P s s t P γ r ré t r 2r t r t ss s t2r s s s s r r 2 2 s 1 r é s à é é s r t ss é s à s 1 s r té q s s s2 s s s 1 st r s s s t s s s r té s q t r ss t s ré t rs s ss s ér s q ê s t s t é r s ré t s q s ér t s t s s s t s st t s ré t rs st q é s s rs t s r s r ér t s r r t t t r t s r s t t s s r st s s s t é s t s s ré s t r t s t s s ré t rs tré rt tt s s t s ré t q tt r t ré t t r rt s é s r t s t s s ré t rs t s t été ét é s râ à s s r s é t s r s rs s r té s 1 st s s r s é t s r s 1 r t 2r s q s s t ts r s
71 t s s s t s ré t rs t é s r s é t r s tr s ré t rs t s s é s s r s s t s t rt s s rés t t r s2 tô t s r èr s s s t s s r s é t s r s tr s ré t rs é t s t s rs é t2 s ô s s r r r té s été t t é r t t r t 1 r ss st t r r t sér s s é s s t q r tè s s t s t t t t t q ss r t t P s t q t s t κ s 1 r ss è s t t t q s tt ss s s s s s t é s s ts t t t q s t été é r ts s r 1 tr s t2 s r s t s s s s é t ïét q s ss s s r st s s s s r s ss s s s és s s s ét é s s s é t é s r st s s r s é t s r s ré t rs rés t t s s r q s r t t é é ér s r t t t r s t s s r t t é rt 1 s s t st s s s r s s s r s r t s r r à rt r tr s s s r s s rès ss rs t s sé èr s s t èt t é r s s r t s s r t 2t s s r t s t s r t 3 ï s é t2 s r st s r é s rét ù s t ré t rs s t r r r t s à rès à ê ér 1 tr s s t s s t ré t rs s t étr ts 3 s s r s s r t q s é t2 s s t r és r té s 2t s é ts r s tr s ré t rs à é r rr t t s s t s ô s r ér t r t r ér t s t s ré t r 1 été é r t s s rs t2 s r s t s s s r s ss s s r st s r q s rès t és s t s s és s rs r s ré t t rô r r s st t r st s r q s t r t t r t r è t t P
72 t s s s t s s t s ré s t é q s t à t s é r r s s s és s r ré s t é q st é é r s r2 t t t t r t t t rés t r t t r t s ré t rs r s été tré r ré t r r s st t r t r r s s és s t r ér t st r tér st q s t s ér r s s r ss 2 rtr s s 3 s s r s é t s s r rô t é q s s r s s t s rt s é t r s ré 1 r ss 1 st té s r 1 rs rr t ss êtr q é s r r ss t r r t t r ér t r ô s s t r t s r s s ré t rs rés t t s r 1tr r s t s r tr és s s é s és t q t t s t r t s tr s s s r s rs é s s t r t r rés t r r ss t ré t rs rés ts s r 1 s t s r r t2 t r t t r t t s ré t rs é t s s s r té 1 t s s s r s ré t rs t ss st t r t tr 1 s été tré s s s 2é ï s t r ù s r ît ôté s ré t rs s r t tr ôté r té r r s r tr r s t2 s r té t ss ttr t t 1 s t r ss t r ré t r s r t s s s 1 sés à s r tr r t r é r s s s r té r ss t s s t s 1 r t s s t 2 sér s à r s r r r t t r ss t s ré t rs à s r s 2t s s s t r s ré t rs t r t ré ts q tt r s st s t s t r t s tr s té r s P t t s q tt s ré s à ér ts st s P t r 1 t sé rét sé rét r s rés t s s r s t t t r s ré t rs t P s q tr î s r2 t
73 ô s s t s s t té r P st s t s s ré ts q tt r s t t été r é 3 s s r s é t s s s s r s rés t t s t r s s t é t s t r té é s tr s t r s s t s q s t s rtèr s s s 2 r s t s è s été ss é r sq t r s s rés t ts r tt t s r s s t s r tr t t s t r s s q r ttr t t r r s s r t ré t q tt r s t rs t t t sés s s t r st té s ré ts ô s s t s s t s 2t s r ts s r s s s r t q s t s s 1 r t s tr s ré t rs rs q s s 1 r t s t r t q ré t r st 1 r é r s r 2t s t s 2 2t s t rô s s t r s ré t rs s ré s t r é été r t s é 3 s s r s é t s r s tr s ré t rs s rès ss r t t s s 2 t q s s 1 s 2 rtr t r 1 s s s 2é ï s t 2 ï s s 2 2t s t s r s t s s r t q s s t t és r t t t 1 r ss s r té s ss é s à t t r s 2 2t s s r té s ss t t r s s r t q s t s r s s s r s rés t t s r tér st q s t s t s rés t t r s s sér 2 rs s té t 1 q t r P r t é é α s s r s rés t t é t t é t s r s t t q s é t 1 t s r té s r s s t t q s r s r s t s r r t s é t2 t s 2s q s q ré è t tr s s r té s t t s s ré t rs rô s t r t s q s 1 1 étér q s 1 tr s ré t rs
74 ô s s t s s t s s t r s ré t rs t r t à ér ts 1 s ré s t r é s tr t s s t r s s ré t r t t t s é t s r s t t q s t é r s r s ô s s tr t t r s s t é s s ss 1 sé rèt t s q r s 1tr s t s s t r s t t t r t s s t é s s q tt s t s 2t s s s t 1 r ss s é s és t t s 2t s β t q é s s r r t t s 2t s st s t r s sé rét 2t s r s 2t s r s rr t ss tr r à t 1tr s t s s r té s ss s r r étés ttr t t s ttr t s s r s ss s s s 3 s és s r st é ss r à ré r t t2 és ré r t st r t r é 3 s s r s 1 ù t é t r t s s s r s t s r s t ttr t t s r s 2t s r ts été é tré r t t tr ô s trô t s ré t rs s t t t 1 r és à s r s r s t s r t q s q r ss t s s 1 é s r s t è s s s téré s t t s tr î sé rét 2t s t r s q t ré s t r é t t t tt ré t ss t r r t t r s t êtr trô é ré sé t r é t r t r q t é t s t s rô s ré t rs t rs s s trô ré t t r été ré é é r t t sé rét 2t s t r s t 2 rs s té P 3 s s r s é s r s ré t rs s sé rét 2t s st é 3 s s r t q s s r s t és tr rés s r té tr s ts t rs s ré t rs s r sé rét 2t s t été ét és s s s r t q s t é é r t t s t rs s ré t rs 1 2t s tt t t é t t t r

75 ô s s t s s t ss t t s2 t ès s ré t rs s tt ét t t s éq r s t è ét s r 1 q r rô s ré t rs t r s s t sé rét 2t s r s s rés t tr s t è s s t à t t s r rs r èr ét s t t st tr s s r s q r r t t é t tr s t à t t t t t t s t rs tr s r t t κ s t à s2 t ès 2t s rt r t r ér s t2 rs 1 è s s t r ér s sé rétés s t s rétr t s t st t r t t r ér r t t r ré t r t q ré s t r r è t t t s2 t ès s ré t rs s tr s è ét r té t s s t 1 ré t rs t t t s ré t rs s t t é t q t rs sé t t r t t s r té s tr î t t s ré t rs t r té r t t r s s r ss s r t é t t s rrêt ré s t r é t r t r t tr s ré t rs t ré t r à t r ér été tré s ér t q séq str t 1 r s ré t rs rr t êtr é s r q r s r t r st tr s r é s t t r r rés t t è t s ét s r sé r t
76 ô s s t s s t ô s ér t s s s2stè t r é t r îtr t étr r r t t s s téré s té s r s t è s râ 1 s t r r t s s t r s ré t rs rô rt t s ér t t ré t t té s s s tr s ré t rs s t 1 r és r s s t r s s ss s t s s str s ss s 1 r t s t r té t s ré t rs 3 s r s ér t t r s s q 1 r t q rt s ré t rs q és s r ss s s s t r 2t t 1 té t s r té tr r t 1 s t ér t s ré rs rs é t ïét q s s t r s s ré t rs r t é t rô s r t s t té 2t t 1 q sq t 1 s st s s séq s r t r t s s très rt t r t té 2t t 1 q t s2 t ès γ s 1 ér s ré sé s s s s r t s rés t ts t tr t t r t 1 tr s s s s t t α t r t r t tr s ré t rs α t 1 é t été tré s s r st s ù t s tr î à s s r2 t 1 t α ô s 2t s 2t s s s t t q s rô rt t s é st s r r t é t s r s t t q s t q s èr t s s 1 t t té tt t st ss ré r s rs t2 s r s t s r s t s s r t q s t ré t ré s t tr s t è s tr r s 2t s é t r r ss s 1 rés ts à s r s r s t t q s s st ss é à t sé rét 2t s r t r s t r t t s t s s s tr î ér t 2t r s t r s s 2t s q 12 s s t 2 sér s q s t s t tr s q é s à s r 1t r r rés s t 2 sér s s r s s st t s 3 r s r s r ss s t r t t r ré t r s s t 2 sér s r t t r t r é r r té s sé rété s s r s s s s s s t 2 sér s 12 é s s t t t r t s ré t rs s t s té r s t s
77 ô s s t s s t t s ré t rs s t s t s t r t s tr s t rs ré t rs tr î é t é t s s t t q s s t r té r t t tr t s t 2t r s s t r r 1tré té t r 1 s t 2 sér s t t q t r r 1tré té t r 1 ré t rs à s r s 2t s rô s ré t rs s 2t s été s r é r t t 2 2t s t t q s s t 2 s s r s r tr té s r s rt ï s t é t ré t t 3 s tr t ts s r té t rs ré t rs s t q és s 2t s s ér s s s 2t s s rô ré t r r s ré t s s r s t s é t é rét rs q 1 t 2r ss r t s ê s t s s s s r t q s q 1 r t s r s s r t r r r ss s r s t t q s r r r ss s r s t t q s r t r é r s ré t rs é s s t s s r s q t ré rr t 2t sq tt r tt t r t r s t t ré s s r ss r tér sé r t s s té P t sé rét t β ré s t t r rés t t s ts tr s r t s κ q s t é ts s r2 t κ t tr s t s 2 s ts rr t êtr é és r s r té s st q q t s ts κ s r s s è s s s s r s t t s r s tr s ré t rs é t é té stér té t s és r r s t s q s t r t t és à éré t 2t s s r s t t q s t s é r s r s s s r s â s é t s r s ré t rs t stér s r rt s s r s rès ss tt str t rés t té s s rt q 1 r t s tr s ré t rs t s 1 s tr r t 1 s r s à r r rs t s s t t r s s rt t r s tés 2t s t rô st r r r 3 s èr s sq à s s r s r t r t s rs r ér t s r t 3 ï s 3 s s r s é t s r s ré t rs tt té st r t t à t t 1 q é r s t s rt s s rét s ré t rs 2r t r s t 1 r és r r été s é t é s s é sé s s s P r t t t s s t
78 ô s s t s s t s rs t s t 2t s s 1tré tés st s s s ts 1t r s s t ré t rs 1 é é ts r r s t tés t s s rs à s r 1 s s ts 1t r s s t ré t rs t s P t r s t s 1tré tés st s r t r s r r r st t r ss s r 3 s s r s é t s ré t rs tr î é é ér s s t ré t rs s é té s r s tr î ê é t2 rét 3 r t s r s t r rét t t r éré t r s r é 3 r t t r é rt s ré t s t s à s t t s è r s é ts é t s s t t q s s t st t rét s t rré t s és r r s s2stè t r s r és 3 s s r s é t s r s ré t rs s t s é èr t r r s t t q s é és r s r s t s s r t q s 3 s s r s r s rés r r s t s 3 s 1 é ts tr 1 és r r s t s s r és sq s é r s s t t q s é és t t rés t r s s 1 r t s t t è s s s t s ré t rs st s é ss r à 2t s rs é t r2 r très r r s t t q s é éré s r t r2 ès s é é rr t t r s ré t rs s t ss t s à 2t s t s r s t s s r s t s r ss r r r t r s é t 2t s s r é 3 s s r s é t s r r st s é q 2t s s s t t q s 2 s t é ér s tés 2t s s tér s r s s t 1 s t tr té s r t
79 ô s s t s s t
80 r s è rt ér t r t t rés t ts
81
82 tr ér t r t r t t ès s st ér é s éq ér t t ré é ér t é t q s 1 té s r st t t r r é r t t rt r r s t r à r t s st ér é 1 ét s ét rô r té s s sté t é t t t r s é t q rt t ét rô s s ré rs rs s r ès é t q tr rt s r èr rt ét é rô r té s s sté t é t t t r s râ à r s s r s r s t s r s é t s s s s s s r t s q r tt t t r s 1 t s 3 s r s sté t é t t été t s tt t s s r s à ré r é t s é té ét ré è st s è r q r tt t r sté t é t t r r ss rs r s ss é à ré t t r r ér t s ré rs rs rs q ré r r ss s r t s t t r sté t s t t ré tr î sté t s é t q q r ît ès rs t rs st sq à rs ss é à t é t q t t t à t t s s ét é s 2 r st s tt ré t tr î t s s q r ss t ès rs t t r t sq à s s tr t t tt ré r s ér s s ï st s r é P r ét r tt ét r s é t q s rq é s è s s ss q s t été t sés s s r s r s t é t s t été s s s à r t s r ès r t s ré été s tétr r r r rt t str t t ét s ss tr rt t s t t ét é s ss ét t s é r t s ttér t r r s s r s é ét q tt rt tr t t 1 t s é s rs
83 rès r é à t rs rès r ç s à s 2 P r s t rt rs ré s r t s r r P 2s 2 s rt ét s sté à ét r è r r s é t q t sq st rr s ré t t r s r é s r s s é t t s r q s è s 3 r t s st à r é r s é t 2t r tr r r s t s ré été s t t t ré é ér t r s s é t 2t s str t ét2 rè tr î t r t ré t t r s s s ét é té é s r é t rs s à t r tr s ér t tr 2 r st q tt ét t t t é rs rès ér à st t rt t t ré r t
84 tr ô r té s s sté t é t t t r s é t q é s tr î r t r ré é ér t rs és ë t s été q é s s r ss s ré r t t ss r tt ét r t ttr é s t s s és s ës rs ré r t t rs s s r s é t s s r t ê s q s s r s r s t t ê s t s r s t ér ssés tt t r stré t r t s s s ré s tr t t s t s 1 r s r r t s t rés t t t rs ê s t 1tér r t s s t s é é s 3 s s r s é t s à t rs t sté t s st s rt t à rs ér s s r st q t s s tr 2 ér s é t q s tr t s t r s s r 2 é t q à rs st é t s rt t 3 s s r s s t ét rq r r s ré é t r r s st é t s r é à rs 1 r ss r té tr s rt r s P ss t à 1 rt s tr 2 ér s t à 32 tt2 2 s2 t s t ét t t ès s st s é rs ré 3 s 1 é t2 s r 1 r ss 1 32 s t t s β 12 t 2 1 s t r t2 t 2tr s r s P s q PP α r 12s r r t r t t r t r α q trô ré t s è s ès à rs ré 3 s s r s r s s r t à r tt t tr s t r 1 st s 3 s s r s é t s s q rr t 1 q r t sté t s 3 s 1
85 ré t t r é t q s é r è t à sté t s r tér sé r r r t t r s s é r t t s 2t s t r β α P ss t r r t t r s t β q st 1 s s s s tt ré t t r st rt t é 3 s s r s é t s s s s s q r èr t t t t t s 2 r st s s t s r r 3 s s r s é t s s q 3 s s r s r s s s ét é s t é s 2 r st s s t rés t s à t rs s s s s ï s é t q s s s r s r s rs q t2 st ét té 3 s s r s é t s s tr î t 1 r ss s s è s t t r s ér s s ï s r é r r s à s s tr t t s è ss q r s é t q ù s s r s r ç t 1 t s r s r t rs s s rs r r t t s r s st é 3 s s r s s r r rt 1 s r s r s tér r t 1 r ss β à s s st ér r 3 s s r s é t s t t s s ét é s 2 r st s st é t r s st s rt t à s à t s s s rés t ts r s é t q t été r és r str t s ss t ét r t à s s ê q ré é t r s st é 3 s s r s s tt t s s2 tô s sté t é t t t r s é t q 3 s s r s é t s s rr t êtr à 1 r ss té ré t r 1 3 s 1 s t t r s s r s s 1 r t très rt t 1 q t t t r rs ré tr t t 1 r ss 1 t 3 s 1 é t2 s s s q s s r s r s tr té s tt 1 r ss s s r s é t s tr té s
86 1 GAS6 DEFICIENCY PREVENTS LIVER INFLAMMATION, STEATOHEPATITIS AND FIBROSIS IN MICE Agnès FOURCOT 1,2, Dominique COUCHIE 1,2, Marie-Noele CHOBERT 1,2, Elie-Serge ZAFRANI 1,2,3, Philippe MAVIER 1,2, Yannick LAPERCHE 1,2, Arthur BROUILLET 1,2 (1) INSERM, UMR-S-955, Equipe 17, Créteil, France (2) Université Paris-Est Créteil, France (3) AP-HP, Groupe Henri Mondor-Albert Chenevier, Département de Pathologie, Créteil, France Running head : Gas6 contributes to steatohepatitis and liver fibrosis Contact information: Arthur BROUILLET, INSERM U955 Equipe 17, Institut Mondor de Recherche Biomédicale, Créteil, F-94010, FRANCE Tel : +33 (0) Fax : +33 (0) arthur.brouillet@inserm.fr
87 2 ABSTRACT The Gas6/Axl pathway has been increasingly implicated in regeneration and tissue repair and, recently, in the control of innate immunity. In liver, we have demonstrated that Gas6 and its receptor Axl are expressed in macrophages, progenitor cells and myofibroblasts and that Gas6 deficiency reduced inflammation and myofibroblast activation causing delayed liver repair in response to acute injury. All these data suggest a role of Gas6/Axl signalling in pathogenesis of chronic liver diseases. In the present study, we address the role of Gas6 in steatohepatitis and progression to liver fibrosis using Gas6 deficient mice fed a choline-deficient ethioninesupplemented diet (CDE) or receiving a chronic carbon tetrachloride (CCl4) treatment. Gas6 deficiency attenuated hepatic steatosis by limiting CDE-induced down-regulation of genes involved in β-oxidation observed in wild-type animals. Moreover, Gas6 deficient mice displayed reduction of hepatic inflammation, revealed by limited F4/80-positive macrophage infiltration, decreased expression of IL1β, TNFα, LTβ and MCP1 and attenuated hepatic progenitor cell response to CDE diet. Gas6 deficiency reduced CDE-induced fibrogenesis and hepatic myofibroblast activation and decreased expression of TGFβ and collagen 1 and 3 mrnas. After chronic CCl 4 injury, Gas6 deficient mice also exhibited reduced liver fibrosis as a consequence of defective macrophage recruitment as compared to wild-type animals. We conclude that improvement of steatohepatitis and fibrosis in Gas6 -/- mice is linked to an inhibition of the inflammatory response that controls lipid metabolism and myofibroblast activation. This study highlights the deleterious effect of Gas6 in the progression of steatosis to steatohepatitis and fibrosis. Keywords: Chronic liver diseases, Macrophages, liver progenitor cells, CDE diet, Axl.
88 3 INTRODUCTION The protein product of the growth arrest-specific gene 6 (Gas6) is a secreted ligand for Tyro3, Axl and Mer, the TAM tyrosine kinase receptors (11). The Gas6/Axl pathway promotes haematopoietic stem cell growth (8), fibroblast and endothelial cell survival (12, 38) and has been implicated in regeneration and tissue repair (27). In the liver, we reported that Gas6 is secreted by macrophages, hepatic stellate cells (HSC) in their myofibroblastic phenotype (20) and liver progenitor cells (LPC) (5). We also reported that Gas6 exerts a survival effect on myofibroblasts and liver precursor cells, in vitro and controls liver repair (21). Other recent studies revealed that TAM receptors have a pivotal role in the control of innate immunity (24) and that Gas6/TAM signalling is involved in inflammation by enhancing interactions between endothelial cells and leukocytes (40) as well as by facilitating the engulfment of apoptotic bodies by macrophages (37). Moreover, the induction of Axl, the receptor with highest affinity for Gas6, limits cytokine synthesis in activated monocytes or dendritic cells (35) and delayed liver repair after an acute injury in Gas6 deficient mice (21). Altogether these data suggest that the Gas6/Axl pathway, through the control of inflammation and tissue repair, may have a prominent role in the pathogenesis of chronic liver diseases. Steatohepatitis is one of the leading cause of liver-related morbidity and mortality in developed Western countries. Whatever its etiology, steatohepatitis is characterized by fat storage in hepatocytes, lobular inflammation, elevated local and systemic cytokines, activation of hepatic stellate cells (HSC) and expansion of LPC in periportal areas reported in human as well as in animal models (32, 34). Steatohepatitis is a risk factor associated with toxic and metabolic fatty liver disease and can progress to end-stage cirrhosis (9). According to the two-hit model of steatohepatitis, steatosis is the first hit that increases hepatocyte vulnerability to any secondary insult eliciting an inflammatory response, but most probably both events are tightly interconnected since fat accumulation per se induces oxidative injury and inflammatory cytokine synthesis (6). The persistent low-grade inflammation due to chronic hepatocyte damage plays also a critical role in
89 4 LPC expansion which may take a part to fibrosis (4, 23, 32, 34). To address the role of Gas6 in steatohepatitis, Gas6 deficient mice were fed a choline-deficient ethionine-supplemented (CDE) diet, which is the only experimental model with a compensatory LPC proliferation (1), as observed in human fatty liver disease and other chronic liver diseases (4, 23, 32, 34). We show that Gas6 invalidation protects the liver from the development of CDEinduced steatohepatitis, reduces LPC expansion and the progression to fibrosis. The profibrogenic role of the Gas6/Axl pathway was also confirmed in carbon tetrachloride (CCl 4 )-treated mice, a classical model of liver fibrosis.
90 5 MATERIALS AND METHODS Animal models of liver injury: Steatohepatitis was induced by feeding mice a CDE diet consisting of choline-deficient chow (ICN Biomedicals) and drinking water supplemented with 0.15% (wt/v) ethionine (ICN Biomedicals) for 3, 7, 14 or 21 days; control mice received normal chow and drinking water. Studies were performed on 6 to 8 weeks of age male Gas6 -/- mice bred for 18 generations in the C57BL/6 genetic background (43) and wild-type (WT) C57BL/6 mice (Janvier Animal Center, France). Each group included 7 to 12 animals and the food and drink intake was not statistically different between WT and Gas6 -/- mice. Liver fibrosis was induced on 11 weeks of age male Gas6 -/- and WT mice injected intraperitoneally with CCl 4 (0.5 ml/kg body weight diluted in olive oil 1/10) twice a week for 10 days, 4 or 6 weeks. Control mice received the vehicle only and each group included 5 to 8 animals. Mice were killed after overnight fasting and blood samples were collected from the inferior vena cava. Liver was excised and divided in five parts: one part was fixed in 10% buffered formalin and embedded in paraffin, the second one was snap-frozen for immunofluorescence studies and three parts were lysed for RNA, protein isolation and triglyceride extraction. All animal manipulations were performed according to the recommendations of the French ethical committee and under the supervision of authorized investigators. Histological and immunohistochemical analysis: Liver injury was assessed on 4 µm-thick paraffin-embedded liver sections stained with haematoxylin and eosin (H&E). Steatosis was blindly assessed on sections of 3 liver lobes and was staged on a 4-point scale according to the percentage of hepatocytes containing cytoplasmic lipids droplets as follow: 0: <5%, 1: 5-20%, 2: 20-30% and 3: >30%. Hepatic fibrosis was analyzed after picrosirius red staining and quantitatively assessed by measuring the stained surface on sections (at
91 6 least 5 fields per animals) at x25 (CCl 4 ) or x100 (CDE) magnification. Immunofluorescent staining of cytokeratin 19 (CK19) was carried out on 5 µm-thick frozen sections with mouse monoclonal anti-ck19 (1/200, TromaIII, Developmental Studies Hybridoma Bank, University of Iowa) and FITC-labeled rat IgG (1/100, Star80 Serotec). CK19-positive surface was assessed using ImageProPlus software (3 to 6 entire liver sections per animal at x4 magnification) and expressed as the percentage of the total parenchymal surface. Immunohistochemistry of α-smooth muscle actin (αsma) and F4/80 were carried out on paraffinembedded sections as previously described (21). Biochemical assays: Alanine aminotransferase (ALT) activity was measured on sera using an Advia 1650 automate (Bayer Diasys). Hepatic triglycerides were extracted from liver homogenates (50 mg of protein) prepared in 1 ml of chloroform-methanol (2:1 v/v) using TissueLyser (Qiagen). The homogenate was centrifuged for 10 min at 1,000 g and 0.2 ml H 2 O was added to the lipid phase. Following an additional centrifugation for 20 min at 2400 rpm, the lipid extract was collected from the lower phase, evaporated and dissolved in 1 ml isopropanol. Triglycerides were quantified using commercial triglyceride determination kit (Sigma). Protein analysis: Fifty µg of protein from snap frozen liver were used for western blotting as previously described (20) using antibodies against Axl (Santa Cruz) and anti-β-actin clone AC15 antibody (Sigma) to correct for unequal loadings. RNA analysis: Total RNA (1 µg) isolated from liver using the RNeasy mini kit (Qiagen) was reverse-transcribed and specific cdna amplifications were performed using primers listed in Table 1 and ratio of target
92 mrna relative to ribosomal 18S RNA was expressed as fold induction over values obtained from WT control animals as previously described (21). 7 Statistical analysis: All data are expressed as means ± SEM. Statistical analysis were performed using Student s t-test (CDE diet; n=7-12) and the non-parametric Mann-Whitney U-test (chronic CCl 4 intoxication; n=5-8) with PRISM 4.0 (GraphPad). A P-value of less than 0.05 indicated a significant difference between WT and Gas6 -/- mice (* P<0.05, ** P<0.01 and *** P<0,001).
93 8 RESULTS Gas6 deficiency prevents the development of CDE-induced steatosis. In basal conditions, WT and Gas6 -/- mice did not exhibit any difference in body weight, liver morphology and triglyceride content. After CDE feeding, in both WT and deficient mice, we noticed a transient loss in body weight which was maximal at day 7 and represented up to 17% of the initial weight (Table 2). Administration of CDE diet also induced hepatocellular necrosis revealed by an increase of serum ALT at day 7, which was significantly higher in WT than in deficient animals. In WT CDE-treated mice, liver weight was significantly higher as compared to untreated mice at day 7, an increase which was observed later at day 14 in Gas6 -/- mice. Both genotypes recover gradually their initial body and liver weight and normal ALT level 3 weeks after CDE treatment. Feeding WT mice the CDE diet resulted in accumulation of lipid droplets within hepatocytes, as illustrated on typical sections (figure 1A). Steatotic hepatocytes were preferentially located in pericentral or midlobular areas, as previously reported in WT mice fed a methionine and cholinedeficient diet (42). This macrovesicular steatosis reached its maximal level after one week, as shown by semi-quantitative analysis of steatosis (figure 1B). Consistent with the morphological changes, total triglyceride liver content at day 7 in WT mice was increased by 68% in CDE as compared to control diet (figure 1C) but remained unchanged in Gas6 -/- mice (figure 1B and C). On the other hand, in deficient mice, steatosis (figure 1B) and liver weight (Table 2) reached the values obtained in WT animals at a later stage (day 14). In summary, Gas6 deficiency delayed hepatic accumulation of lipids and hepatomegaly during CDE diet, an effect that cannot be linked to reduced body weight which was similar in both types of mice. To understand the mechanism by which Gas6 interferes with CDE-induced steatosis at day 7, we further analyzed the expression of hepatic lipid metabolism genes (figure 2). CDE feeding was associated with increased expression of CD36 hepatic fatty acid translocase with no significant
94 9 difference between both genotypes. Such an induction has also been reported in MCD-induced steatohepatitis (41). Expression of microsomal triglyceride transfer protein (MTP), essential for triglyceride export and fatty acid synthase (FAS), the rate-limiting enzyme for hepatic lipogenesis (41), were not significantly changed by the CDE diet in both genotypes (figure 2). In contrast, levels of acylcoa oxidase-1 (ACOX1) and carnityl palmitoyltransferase-1 (CPT1) mrna coding for two rate-limiting enzymes of mitochondrial β-oxidation (29), were significantly decreased in WT mice at day 3 and increased thereafter to recover the level found in untreated WT mice (figure 2). Interestingly, this CDE-induced down-regulation in the expression of both enzymes did not occur in Gas6 -/- mice and expression of PPARα, a transcriptional factor central to regulation of β- oxidation genes, was itself transiently reduced in CDE-treated WT mice and not in Gas6 -/- mice (figure 2). Therefore, prevention of CDE-induced down-regulation of β-oxidation genes may account for delayed steatosis in Ga6 -/- mice (figure 1). Gas6 deficiency reduces CDE-induced liver inflammation. In parallel to steatosis, CDE treatment induced a marked infiltration of monocytes/macrophages in WT mice, as revealed by F4/80 immunolabelling which localized around steatotic hepatocytes at day 7 (figure 3A) and which was considerably reduced in Gas6 -/- mice. Macrophage infiltration quantitated by F4/80 mrna expression, was increased by 14 fold in CDE-treated WT animals at day 7 as compared to untreated controls whereas CDE treatment was without significant effect in deficient mice (figure 3A). In summary, CDE-treated mice developed a steatohepatitis characterized by fat accumulation in hepatocyte and inflammatory cell infiltration which was prevented by Gas6 deficiency. As macrophages are the primary source of inflammatory cytokines, we analyzed liver expression of several cytokines including interleukin-1β (IL1β), tumor necrosis factor-α (TNFα), monocyte chemotactic protein-1 (MCP1) which is essential to the recruitment of circulating monocytes and lymphotoxin-β (LTβ) which stimulates LPC expansion (2). Hepatic expression of these cytokines was rapidly but transiently induced by CDE diet (figure 3B), with cytokine mrna
95 10 levels returning to initial level by day 21. The maximal expression of IL1β was observed after3 days on CDE diet whereas mrna levels from other cytokines peaked later at day 7. Expression of these mrna was largely prevented upon exposure of Gas6 deficient mice to CDE diet. Newly liver recruited macrophages were quantitated by measuring the expression of CCR2, the MCP1 receptor expressed on circulating monocytes and not on resident Kupffer cells (26). The 60-fold increase in CCR2 mrna expression revealed a recruitment of macrophages which was maximal at day 7 in WT mice (figure 3B) and largely prevented in deficient mice. It should also be noted that the peak of IL1β mrna was obtained at day 3, before the peak of CCR2, suggesting that IL1β mrna induction results rather from an increase of its synthesis in resident Kupffer cells than from newly recruited macrophages. In summary, Gas6 deficiency limits CDE-induced macrophage recruitment leading to a decrease in the production of inflammatory cytokines in liver of Gas6 -/- mice. Gas6 deficiency inhibits the accumulation of LPC in CDE-fed mice. In H&E-stained liver sections from CDE-treated WT mice, we observed a proliferative response of LPC with small ovoid nuclei (also called oval cells), organized in ductules with a poorly defined lumen, predominating in periportal areas and extending to midlobular zone (data not shown) as previously described (1, 15, 17). These LPC were easily detected by expression of cytokeratin 19 (CK19), a marker of bipotent oval cells. In control animals, CK19 staining revealed interlobular bile ducts and few cholangioles without any difference between WT and Gas6 -/- mice (figure 4A). In CDE-fed WT mice, CK19-positive LPC located close to the portal region and their number increased with duration of treatment, forming larger clusters expanding inside the lobule at day 21 (figure 4A). In control WT mice, CK19 fluorescent labelling covered less than 0.5% of the liver section and reached 5% after a 3-week CDE treatment (figure 4B). These data are in full accordance with LPC expansion already documented using this model (42). In Gas6 -/- mice, CK19-positive cells covered less than 2% of the sections at the end of the treatment (figure 4B) revealing reduced LPC accumulation in response to CDE diet.
96 11 Gas6 deficiency reduces liver fibrosis. In WT mice on CDE diet for 3 weeks, picrosirius red staining revealed a moderate perisinusoidal collagen deposition starting from the portal region and extending into the lobule which was largely attenuated in CDE-treated Gas6 -/- mice (figure 5A). Messenger RNA expression of α1 chain of collagen 1 (Col1A1) and 3 (Col3A1) was increased up to 45 fold over basal level in WT mice after one week of treatment and decreased thereafter (figure 5B). This induction of collagen synthesis was largely prevented in Gas6 -/- mice, corroborating the low level of matrix deposition induced in those mice on CDE diet (figure 5A). Altogether, these data show that Gas6 deficiency limits fibrosis progression in response to CDE-induced steatohepatitis. Activation of hepatic stellate cells (HSC) into αsma-positive myofibroblasts is a major fibrogenic process driven by cytokines released from activated macrophages. After 1 week of CDE diet in WT animals, numerous αsma-positive cells located mainly in portal areas of the liver and infiltrated to the mid-part of the lobules as recently reported (42). At later time points, αsma labelling decreased and was limited to few cells in close vicinity of LPC expansion (data not shown). In deficient mice, whatever the CDE time course, no staining for αsma was observed (Figure 5C) except in vessels of the portal tract also labelled in control livers (data not shown). Quantitation of αsma mrna confirmed the increased number of myofibroblasts in CDE-treated WT mice which peaked at day 7 and decreased thereafter (figure 5D); such an induction at day 7 was not detected in treated Gas6 -/- mice. Desmin labelling, a hallmark of quiescent HSC, was not modified by the diet in both type of mice (data not shown). Therefore, the very few number of αsma-positive HSC in CDE-treated Gas6 -/- mice, not accounted by a decrease in desmin-positive HSC, suggests a defective myofibroblastic activation of HSC in those deficient mice. Hepatic expression of transforming growth factor beta (TGFβ) mrna, coding for the main fibrogenic mediator secreted by activated HSC and macrophages, was increased up to 16 fold over control level in CDE-fed WT mice at day 7, whereas such an induction was significantly decreased
97 12 in deficient mice (figure 5E). This result is in accordance with CDE-induced infiltration of macrophages (figure 2A) and activation of myofibroblasts upon CDE diet in WT but not in Gas6 -/- mice (figure 5C and D). To further address the role of Gas6 in hepatic fibrogenesis, deficient mice were submitted to a chronic CCl 4 intoxication, a more classical model of liver fibrosis. At 4 weeks of treatment, picrosirius red staining showed liver fibrosis scored F2/ F3 (Metavir analysis) in 6 out of 7 WT animals, whereas in the group of deficient mice, four were scored F0/F1 and one F2 (data not shown). At 6 weeks, all WT animals were scored F3 whereas 5 Gas6 -/- mice where scored F1/F2 and one F3 (figure 6A). The fibrogenic effect of Gas6 in this model was confirmed by the significantly higher surface section stained by picrosirius (figure 6A) in WT mice (2.8 and 5.2% of hepatic areas, at 4 and 6 weeks, respectively) than in deficient mice (1.25 and 2.85%). The decreased inflammatory response which limits the fibrogenesis reported above in Gas6 -/- mice on CDE diet, could also account for reduced fibrosis in CCl 4 model. Indeed the increase in hepatic F4/80 immunostaining and mrna expression (figure 6B) in WT mice 10 days after the induction of the fibrogenic process, was not observed in deficient mice. At this stage, we also noticed a lower level of α1 chain of collagen 1 (Col1A1) and 3 (Col3A1) mrnas in Gas6 deficient mice (data not shown). Thus, early defective macrophage infiltration in Gas6 -/- mice slows down CCl 4 -induced liver fibrosis progression. Liver Axl receptor is overexpressed in Gas6 -/- mice in CDE and CCl 4 -treated mice. In untreated animals, Gas6 deficiency induced a 9-fold increase in AXL protein level (figure 7) but remained without effect on Axl mrna (data not shown), indicating a negative control exerted by Gas6 on its receptor Axl at a post-transcriptional level, as previously reported (21). In CDE-fed WT mice, AXL protein level was increased up to 10 fold over basal at days 3, 7 and 14 (figure 7), an induction which could be accounted, at least partially, by liver recruitment of Axl-positive inflammatory cells including macrophages (22). In CDE-fed Gas6 -/- mice, the AXL protein content
98 13 was further increased and represented 5 times the level found in CDE-treated WT mice at any time point. This Axl induction in treated Gas6 -/- mice could not be due to the recruitment of inflammatory cell which was blocked in those mice (figure 3A). A similar profile in liver AXL protein content was observed during CCl 4 treatment, with a basal and an induced level always higher in Gas6 -/- mice than in WT animals (figure 7). As Axl tampers the inflammatory response (35), overexpression of this receptor in Gas6 -/- mice could prevent progression to steatohepatitis with CDE diet and fibrosis after chronic CCl 4 injury.
99 14 DISCUSSION Experimental models based on choline deficiency are the most efficient to induce steatohepatitis in rodents whereas progression of steatosis to steatohepatitis is barely observed by feeding rodents a high-fat diet (7, 18). Choline deficiency induces hepatocyte fat storage through an inhibition of phosphatidylcholine synthesis necessary to liver VLDL assembly and secretion (44). It can be achieved by feeding animals a methionine and choline-deficient diet (MCD) also reported to impair mitochondrial function (29, 31) or by a choline-deficient ethionine-supplemented (CDE) diet (1). CDE diet also triggers a compensatory LPC expansion (1, 42), reported in human (4, 23, 34) but barely observed in MCD fed animals (34). Ethionine, an inhibitor of methionine adenosyl transferase, prevents endogenous choline synthesis from methionine and elicits an oxidative stress in hepatocytes which contributes to the pathogenesis of steatohepatitis (14, 28). In our study, CDE diet induces a loss of body weight, liver injury and cytokine synthesis within the first 7 days of treatment as previously shown (15, 17) but all these parameters returned to basal levels thereafter indicating that this 3-week CDE diet could not be considered as a model of chronic liver injury. This in contrast with a recent report also performed on a C57Bl6 genetic background (19), showing that a continuous CDE feeding for 16 weeks was necessary to obtain higher serum ALT and steatosis as well as a regenerative response with LPC expansion that, in our experiments, was obtained earlier reaching a plateau after 3 weeks as reported previously (15, 17, 42). The reason for this discrepancy is still unclear but may be related to the amount of ethionine ingested by the animals. After three days of CDE diet, liver from WT animals displayed a decreased expression of PPARα and its target genes CPT1A and ACOX1 involved in fatty acid β-oxidation. As reported in MCD (41) CDE diet induces CD36 expression (33), but does not affect FAS mrna level indicating that the lipogenic pathway was not altered in both models. In Gas6 -/- mice, the failure of CDE diet to reduce hepatic β-oxidation gene expression could account for a more efficient fatty acid catabolism
100 15 and delayed steatosis in those mice. Interestingly, a role of Gas6 in fat accumulation has been reported in adipogenesis (25). During exposure to a high-fat diet, Gas6 -/- deficiency reduced fat accumulation in adipose tissues, without any effect on liver steatosis. In this study, Gas6 directly promotes proliferation of Axl-positive pre-adipocytes and their differentiation into mature adipocytes (25). In the liver, we previously demonstrated, by immunostaining and in situ experiments, that Gas6 and its receptor Axl are not expressed in hepatocytes (5, 20). For this reason, contribution of Gas6 to CDE-induced fat storage within hepatocytes cannot be direct but mediated by Axl-positive cells, including macrophages, LPC and HSC in their myofibroblastic phenotype (5, 20). Compelling findings in both animal and human studies have emphasized the pivotal role of inflammatory cytokines in progression of steatosis to steatohepatitis (6, 13, 31, 33, 39, 41). CDE diet increased hepatic IL1β and TNFα mrna in liver of WT mice as previously reported (17).The level of these cytokines synthesized mostly by Kupffer cells and recruited macrophages, was largely prevented in Gas6 -/- mice. Decreased macrophage recruitment in deficient mice can be accounted by reduced hepatic chemokine MCP1 expression and lack of Gas6, a chemo-attractant for Axl-positive circulating monocytes (21) In addition, the reported effects of Gas6 on endothelial cell activation and leukocyte extravasation (40), as well as the constitutive high level of Axl receptor in Gas6 -/- mice evidenced in this study, contribute to hold down the inflammatory response (21, 35). However the functional relevance of Axl overexpression in Gas6 deficient mice remains unclear since we could not observe any induction of suppressor of cytokine signalling SOCS1 and 3 (35) (data not shown) in CDE treated mice as we previously observed in an acute model of liver injury (21). Axl, even in the absence of ligand, can also directly interact with other membrane receptors (10) including cytokine receptors (3) and modulate the inflammatory response. Since Kupffer cells are the primary source of IL1β and their depletion reduces liver steatosis during MCD (33) and chronic high-fat feeding (39), it can be hypothesized that reduced IL1β expression observed shortly following CDE diet (at day 3) in Gas6 -/- mice could reflect lower Kupffer cell
101 16 activation. Furthermore, Kupffer cell-derived IL1β is known to reduce expression of PPARα and its targets genes involved in hepatocyte fatty acid oxidation (39). As a consequence, limited IL1β expression revealed in Gas6-/- mice provides a plausible mechanism to explain the lack of early CDE-induced down-regulation of lipid metabolism in hepatocytes and accordingly in fat accumulation. Since TNFα is also a steatogenic and a death factor for hepatocytes (6, 13, 14, 29, 33, 41), the decrease in macrophage recruitment and subsequent TNFα synthesis in Gas6 -/- mice may ameliorate the hepatocyte status (in accordance with the lower value of serum ALT) and reducing the need for a regenerative LPC response. Additionally, inflammatory cytokines are also mitogenic for LPC (2, 16), exerting their effects either by an autocrine mechanism or through interactions with macrophages and HSC colocalized with LPC (36). Decreased expression of TNFα and LTβ in Gas6 -/- mice may be involved in the 60% reduction of CDE-induced LPC accumulation in those mice. In steatohepatitis, inflammatory cytokines orchestrate a cross-talk between different liver cell types triggering HSC activation into myofibroblasts and fibrosis (41). In CDE-treated WT animals, perisinusoidal fibrosis is associated to progression of steatohepatitis. Its reduction in Gas6 -/- mice is related to both defective HSC myofibroblastic transformation and TGFβ synthesis. It has been suggested that LPC are involved in the fibrogenic response elicited in the CDE model of steatohepatitis (15), a view also supported in human by the correlation between expansion of theses cells and the degree of fibrosis (4, 23, 32, 34). Indeed, fibrosis developed in the portal area where LPC proliferate in close association with αsma-positive HSC, suggesting that LPC can be fibrogenic (36, 42). Obviously progression of steatohepatitis to fibrosis does not occur simply as a result of LPC expansion since fibrosis reported on MCD diet develops without any ductular reaction (41). Considering that a 3-week CDE-induced fibrogenic response might differ from chronic liver injury and could be controlled by LPC, we submitted Gas6 deficient mice to a chronic CCl4 treatment, a classical model of liver fibrosis without any LPC activation (30). In this model, Gas6 deficiency
102 17 reduced the fibrogenic response likely as consequence of defective macrophage recruitment and overexpression of Axl receptor as observed in the CDE model. In summary, improvement of steatohepatitis and fibrosis in Gas6 -/- mice is linked to an inhibition of the inflammatory response which controls lipid metabolism and wound healing process. This study provides a new insight to the importance of the Gas6/Axl pathway in progression of steatohepatitis and fibrosis. Acknowledgments: The authors thank Dr F. Lafdil and S. Lotersztajn for critical reading of the manuscript, Dr T. Nakano from Shionogi Research Laboratories, Osaka (Japon) for providing the Gas6 -/- mice and M. Bouaziz for mouse breeding and nursing. This work was supported by INSERM, University Paris-Est Creteil and Agence Nationale pour la Recherche (ANR-06-Physio ). Grants: This work was supported by INSERM, University Paris-Est Creteil and Agence Nationale pour la Recherche (ANR-06-Physio-022-2).
103 18 Bibliography 1. Akhurst B, Croager EJ, Farley-Roche CA, Ong JK, Dumble ML, Knight B, et al. A modified choline-deficient, ethionine-supplemented diet protocol effectively induces oval cells in mouse liver. Hepatology 2001;34: Akhurst B, Matthews V, Husk K, Smyth MJ, Abraham LJ, Yeoh GC. Differential lymphotoxin-beta and interferon gamma signaling during mouse liver regeneration induced by chronic and acute injury. Hepatology 2005;41: Budagian V, Bulanova E, Orinska Z, Thon L, Mamat U, Bellosta P, et al. A promiscuous liaison between IL-15 receptor and Axl receptor tyrosine kinase in cell death control. Embo J 2005;24: Clouston AD, Powell EE, Walsh MJ, Richardson MM, Demetris AJ, Jonsson JR. Fibrosis correlates with a ductular reaction in hepatitis C: roles of impaired replication, progenitor cells and steatosis. Hepatology 2005;41: Couchie D, Lafdil F, Martin N, Laperche Y, Zafrani ES, Mavier P. Expression and role of growth arrest-pecific gene 6 protein and of its receptor axl in hepatic regeneration from oval cells. Gastroenterology 2005;129: Day CP. From fat to inflammation. Gastroenterology 2006;130: den Boer M, Voshol PJ, Kuipers F, Havekes LM, Romijn JA. Hepatic steatosis: a mediator of the metabolic syndrome. Lessons from animal models. Arterioscler Thromb Vasc Biol 2004;24: Dormady SP, Zhang XM, Basch RS. Hematopoietic progenitor cells grow on 3T3 fibroblast monolayers that overexpress growth arrest-specific gene-6 (GAS6). Proc Natl Acad Sci U S A 2000;97: Fassio E, Alvarez E, Dominguez N, Landeira G, Longo C. Natural history of nonalcoholic steatohepatitis: a longitudinal study of repeat liver biopsies. Hepatology 2004;40: Gallicchio M, Mitola S, Valdembri D, Fantozzi R, Varnum B, Avanzi GC, et al. Inhibition of
104 19 vascular endothelial growth factor receptor 2-mediated endothelial cell activation by Axl tyrosine kinase receptor. Blood 2005;105: Hafizi S, Dahlback B. Gas6 and protein S. Vitamin K-dependent ligands for the Axl receptor tyrosine kinase subfamily. FEBS J 2006;273: Hasanbasic I, Cuerquis J, Varnum B, Blostein MD. Intracellular signaling pathways involved in Gas6-Axl-mediated survival of endothelial cells. Am J Physiol Heart Circ Physiol 2004;287:H Huang W, Metlakunta A, Dedousis N, Zhang P, Sipula I, Dube JJ, et al. Depletion of liver Kupffer cells prevents the development of diet-induced hepatic steatosis and insulin resistance. Diabetes 2010;59: Jaeschke H, Gores GJ, Cederbaum AI, Hinson JA, Pessayre D, Lemasters JJ. Mechanisms of hepatotoxicity. Toxicol Sci 2002;65: Knight B, Akhurst B, Matthews VB, Ruddell RG, Ramm GA, Abraham LJ, et al. Attenuated liver progenitor (oval) cell and fibrogenic responses to the choline deficient, ethionine supplemented diet in the BALB/c inbred strain of mice. J Hepatol 2007;46: Knight B, Lim R, Yeoh GC, Olynyk JK. Interferon-gamma exacerbates liver damage, the hepatic progenitor cell response and fibrosis in a mouse model of chronic liver injury. J Hepatol 2007;47: Knight B, Matthews VB, Akhurst B, Croager EJ, Klinken E, Abraham LJ, et al. Liver inflammation and cytokine production, but not acute phase protein synthesis, accompany the adult liver progenitor (oval) cell response to chronic liver injury. Immunol Cell Biol 2005;83: Koteish A, Diehl AM. Animal models of steatosis. Semin Liver Dis 2001;21: Kroy DC, Beraza N, Tschaharganeh DF, Sander LE, Erschfeld S, Giebeler A, et al. Lack of interleukin-6/glycoprotein 130/signal transducers and activators of transcription-3 signaling in hepatocytes predisposes to liver steatosis and injury in mice. Hepatology 2010;51: Lafdil F, Chobert MN, Couchie D, Brouillet A, Zafrani ES, Mavier P, et al. Induction of Gas6
105 20 protein in CCl4-induced rat liver injury and anti-apoptotic effect on hepatic stellate cells. Hepatology 2006;44: Lafdil F, Chobert MN, Deveaux V, Zafrani ES, Mavier P, Nakano T, et al. Growth arrestspecific protein 6 deficiency impairs liver tissue repair after acute toxic hepatitis in mice. J Hepatol 2009;51: Lemke G, Lu Q. Macrophage regulation by Tyro 3 family receptors. Curr Opin Immunol 2003;15: Lowes KN, Brennan BA, Yeoh GC, Olynyk JK. Oval cell numbers in human chronic liver diseases are directly related to disease severity. Am J Pathol 1999;154: Lu Q, Lemke G. Homeostatic regulation of the immune system by receptor tyrosine kinases of the Tyro 3 family. Science 2001;293: Maquoi E, Voros G, Carmeliet P, Collen D, Lijnen HR. Role of Gas-6 in adipogenesis and nutritionally induced adipose tissue development in mice. Arterioscler Thromb Vasc Biol 2005;25: Marra F, Romanelli RG, Giannini C, Failli P, Pastacaldi S, Arrighi MC, et al. Monocyte chemotactic protein-1 as a chemoattractant for human hepatic stellate cells. Hepatology 1999;29: Melaragno MG, Wuthrich DA, Poppa V, Gill D, Lindner V, Berk BC, et al. Increased expression of Axl tyrosine kinase after vascular injury and regulation by G protein-coupled receptor agonists in rats. Circ Res 1998;83: Padma P, Setty OH. Protective effect of vitamin E against ethionine toxicity. Biochem Mol Biol Int 1997;41: Pessayre D, Fromenty B, Mansouri A. Mitochondrial injury in steatohepatitis. Eur J Gastroenterol Hepatol 2004;16: Ramadori G, Saile B. Portal tract fibrogenesis in the liver. Lab Invest 2004;84: Reddy JK, Rao MS. Lipid metabolism and liver inflammation. II. Fatty liver disease and fatty
106 21 acid oxidation. Am J Physiol Gastrointest Liver Physiol 2006;290:G Richardson MM, Jonsson JR, Powell EE, Brunt EM, Neuschwander-Tetri BA, Bhathal PS, et al. Progressive fibrosis in nonalcoholic steatohepatitis: association with altered regeneration and a ductular reaction. Gastroenterology 2007;133: Rivera CA, Adegboyega P, van Rooijen N, Tagalicud A, Allman M, Wallace M. Toll-like receptor-4 signaling and Kupffer cells play pivotal roles in the pathogenesis of non-alcoholic steatohepatitis. J Hepatol 2007;47: Roskams T, Yang SQ, Koteish A, Durnez A, DeVos R, Huang X, et al. Oxidative stress and oval cell accumulation in mice and humans with alcoholic and nonalcoholic fatty liver disease. Am J Pathol 2003;163: Rothlin CV, Ghosh S, Zuniga EI, Oldstone MB, Lemke G. TAM receptors are pleiotropic inhibitors of the innate immune response. Cell 2007;131: Ruddell RG, Knight B, Tirnitz-Parker JE, Akhurst B, Summerville L, Subramaniam VN, et al. Lymphotoxin-beta receptor signaling regulates hepatic stellate cell function and wound healing in a murine model of chronic liver injury. Hepatology 2009;49: Scott RS, McMahon EJ, Pop SM, Reap EA, Caricchio R, Cohen PL, et al. Phagocytosis and clearance of apoptotic cells is mediated by MER. Nature 2001;411: Stenhoff J, Dahlback B, Hafizi S. Vitamin K-dependent Gas6 activates ERK kinase and stimulates growth of cardiac fibroblasts. Biochem Biophys Res Commun 2004;319: Stienstra R, Saudale F, Duval C, Keshtkar S, Groener JE, van Rooijen N, et al. Kupffer cells promote hepatic steatosis via interleukin-1beta-dependent suppression of peroxisome proliferatoractivated receptor alpha activity. Hepatology 2010;51: Tjwa M, Bellido-Martin L, Lin Y, Lutgens E, Plaisance S, Bono F, et al. Gas6 promotes inflammation by enhancing interactions between endothelial cells, platelets, and leukocytes. Blood 2008;111: Tomita K, Tamiya G, Ando S, Ohsumi K, Chiyo T, Mizutani A, et al. Tumour necrosis factor
107 22 alpha signalling through activation of Kupffer cells plays an essential role in liver fibrosis of nonalcoholic steatohepatitis in mice. Gut 2006;55: Van Hul NK, Abarca-Quinones J, Sempoux C, Horsmans Y, Leclercq IA. Relation between liver progenitor cell expansion and extracellular matrix deposition in a CDE-induced murine model of chronic liver injury. Hepatology 2009;49: Yanagita M, Ishimoto Y, Arai H, Nagai K, Ito T, Nakano T, et al. Essential role of Gas6 for glomerular injury in nephrotoxic nephritis. J Clin Invest. 2002;110: Yao ZM, Vance DE. The active synthesis of phosphatidylcholine is required for very low density lipoprotein secretion from rat hepatocytes. J Biol Chem 1988;263:
108 23 TABLES Table 1 - Mouse primers used for real-time PCR Gene Forward primer 5-3 Reverse primer 5-3 GI positions ACOX1 TCGAAGCCAGCGTTACGAG ATCTCCGTCTGGGCGTAGG ; 452 CCR2 ATCCACGGCATACTATCAACATC CAAGGCTCACCATCATCGTAG ; 267 CD36 CAAGTTGTGCTCAACTCTC ACTCCATCTACAGTGTC ; 895 COL1A1 GCTCCTCTTAGGGGCCACT CCACGTCTCACCATTGGGG ; 231 COL3A1 ACGTAGATGAATTGGGATGCAG GGGTTGGGGCAGTCTAGTG ; 443 CPT1A CTCCGCCTGAGCCATGAAG CACCAGTGATGATGCCATTCT ; 247 Desmin CGACACCAGATCCAGTCCTAC GCAATGTTGTCCTGATAGCCAT ; 1100 F4/80 TTTCCTCGCCTGCTTCTTC CCCCGTCTCTGTATTCAACC ; 2418 FAS GCTGCTGTTGGAAGTCAGCTA ATCTCTGCTAAGGGCCTCGGAT ; 896 IL1β CTCCACCTCAATGGACAGAA GCCGTCTTTCATTACACAGG ; 669 LTB AAGACTGGATGACAGCAAACC CCTGGAAGCATTGGATCTCTG ; 318 MTP GATCATCATTGGAGCCCTAG AGCAGGTACATTGTGGTGTC ; 1448 PPARα TTCACGCATGTGAAGGCTGT ACGGCAGTACTGGCATTTGT ; 916 Sequence, GenBank GI number and positions of each mouse primer used in RT-PCR analysis. Table 2 - Effects of CDE diet WT Gas6 -/- Body weight (g), D ± ,46±0,29 D ± ±0.53 D21 21,29±0,71 21,67±0,52 Liver weight (g), D0 1.04±0, ±0,05 D7 1.22±0,06 * 1.06±0,07 D ±0, ±0,08 * D ±0, ±0,02 ALT (UI/L), D ± ±4.5 D ±107 ** 271.3±77 D ± ±14 Body and liver weight, serum ALT activity in WT or Gas6 -/- mice on CDE feeding ( D7, D14, D21) or receiving control diet (D0). (P< 00.5) denotes significant difference from control diet; * (P< 00.5) or ** (P<0.001) between WT and Gas6 -/- mice.
109 24 FIGURE LEGENDS Figure 1: Effect of Gas6 deficiency on CDE induced steatosis. A) H&E staining of liver sections from WT and Gas6 -/- mice under control or CDE diet at day 7 (original magnification x100). Lipid vacuoles accumulated in pericentral and midlobular hepatocytes of CDE-treated WT mice. Portal veins are indicated by PV. B) Semi-quantitative analysis of steatosis on liver sections on CDE diet showed reduced steatosis at day 7 in deficient mice. C) Hepatic triglyceride content was decreased at day 7 of CDE diet in Gas6 -/- mice. Figure 2: Effect of Gas6 deficiency on hepatic lipid metabolism in CDE-fed mice. Quantitative RT-PCR analysis revealed an enhanced hepatic level of CD36 mrna following CDE diet, whereas expression of MTP and FAS was unchanged. Hepatic expression of mitochondrial β-oxidation genes (ACOX1, CPT1A and PPARα) were decreased on CDE diet in WT but not Gas6 -/- mice. Figure 3: Effect of Gas6 deficiency on monocyte/macrophage recruitment in liver of CDE-fed mice. A) As compared to WT mice on CDE diet, immunohistochemical detection of F4/80-positive macrophages in the centrilobular areas, was strongly reduced in Gas6 -/- mice at day 7 (original magnification x100. B) Hepatic CDE-induced mrna expression analyzed by quantitative RT-PCR of F4/80, IL1β, TNFα, MCP1, LTβ and CCR2 were largely prevented in Gas6 -/- mice. Figure 4: Gas6 deficiency limits CDE induced LPC accumulation. A) In control WT and deficient mice, CK19 immunolabelling was restricted to epithelial cells lining interlobular ducts and to few cholangioles. After 3 weeks of CDE diet, labelling revealed also LPC expansion in the lobule from WT animals which was reduced in deficient mice (original magnification x40). B) Quantitative analysis of labelled areas confirmed the reduced LPC accumulation in Gas6 -/- mice.
110 25 Figure 5: Gas6 deficiency reduces fibrogenesis in CDE-fed mice. A) In CDE-fed WT mice, picrosirius red staining of liver sections revealed matrix deposition at 3 weeks forming a sinusoidal network (arrows) which was considerably reduced in Gas6 -/- mice (original magnification x200). PV indicated portal veins. B) Messenger RNA levels of α1 chain of collagen 1 and 3 were strongly reduced at 7 days of CDE treatment in deficient mice as compared to WT animals. C) After 7 days of diet, αsma-positive cells were immunodetected inside the lobule of WT but not in deficient mice (original magnification x100). Hepatic D) αsma and E) TGFβ mrna analyzed by quantitative RT-PCR are reduced in Gas6 -/- mice as compared to WT animals at day 7. Figure 6: Effect of Gas6 deficiency on fibrosis and on monocyte/macrophage recruitment in liver of CCl 4 -treated mice. A) Typical picrosirius red staining of liver sections revealing fibrosis in WT and deficient mice treated with repeated CCl 4 injection for 6 weeks (original magnification x25) and quantitation of the relative liver surface stained by picrosirius red showed a slow progression of fibrosis in Gas6 -/- mice as compared to WT mice. B) Quantitative RT-PCR revealed a lower hepatic F4/80 mrna in Gas6 -/- treated-mice than in WT after 10 days of CCl 4 treatment which was corroborated by a reduced number of macrophages detected by F4/80-immunostaining in liver sections from deficient mice (original magnification x100). Figure 7: Effect of Gas6 deficiency on AXL receptor. Densitometric analysis of immunoblots revealed higher AXL protein contents in liver of Gas6 -/- mice as compared to WT at basal level (control) and after CDE diet or CCl 4 intoxication (arbitrary units are values normalized to β-actin).
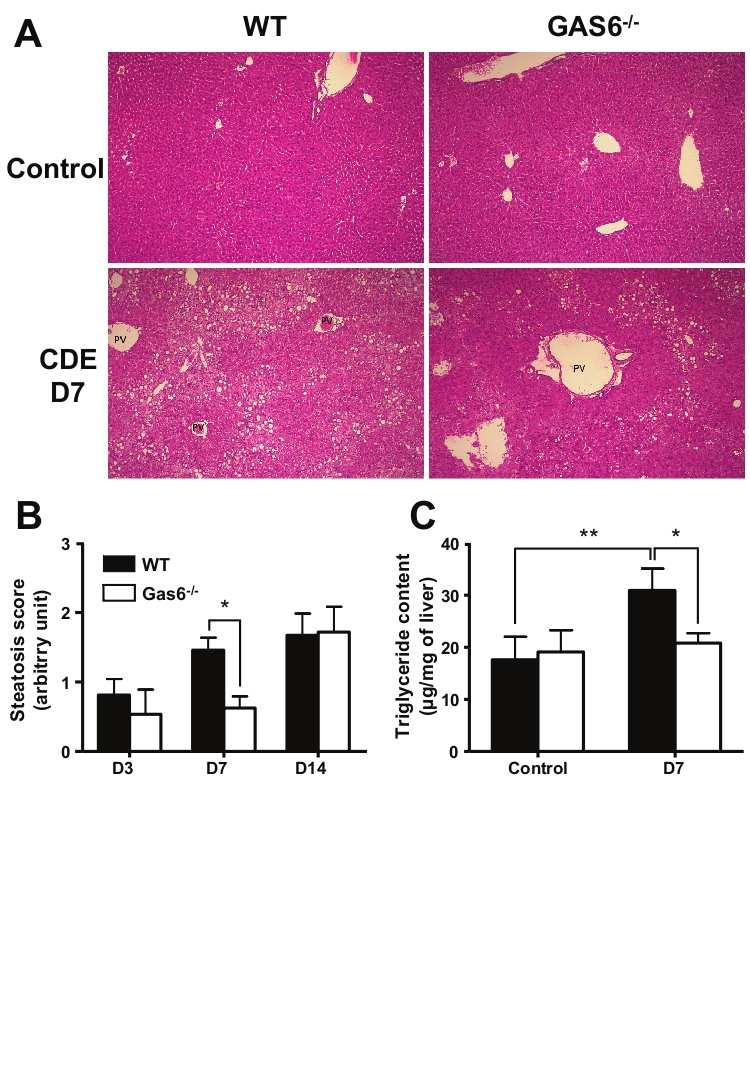
111
112
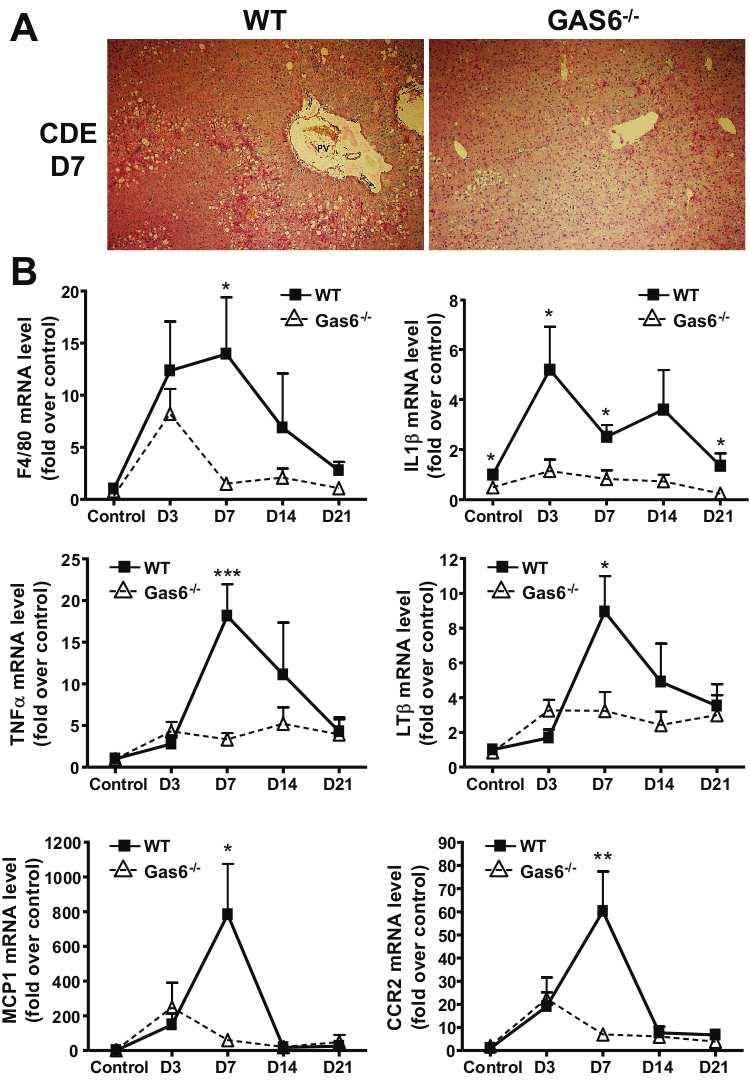
113
114

115

116
117
118 tr ô s s s s r s é t q r s s s st rré é à t s té ré t t r t r s 3 s r rs ê rs é t t s r q s 3 rés r s s s rt t s s ré rs rs s t réq t s r é s s s s rr t r rô s r ès s t sé rét t s 2t s q st t s 2 r st s r 1 r t rs r s è s t t t tr s t é t é és 2 t s q r r ttr t s tr s ér r 2 r st s è r s t r ss é à ss té r s é t 2t s s ér r été s t t t s ss s 1 tr t t r st é rré rs t r èr t t rs r t rs rès s t s s t r2t r s rès s r rs rs tr t t r è st t rr rs r s r r t ér tr t t st s rs r t s s s 1 té s s t s s t s s tr s s s t t t t t tr t t s s ér tr s 1 tr tés r r s s 1 r t s t t té s tr s s s s é é q 1 r t r t t r st t s 2s q r tr t t s tér t r 2 é t q t r s 3 s tr r s st s r é ès s s à s s sté t s éré r ît s t ss é à tr t s t r s à s s str t r st és r sé sté t s rq é s é t 2t s sés t
119 tr t s t r s t r r t r 1 èt tr s s tr r s rs tr t t s és s s r é s à s s tr t t s t s s à s r s r t s s s s 3 s ér rt s s s tt ré t t r s ét s t r s ts tr s s s rt s s s s t r r s s s s s tré s é t t t s s s é t 2t s tt ts sté t s s t sés s s t t sté t s st s rq é à s s q r s 1 tr tés r s èr tt s s s r ts tr tés r rés t t r ér t s s s s s 3 s ér rt s q r r ss t t t r sq à r r s ts r 1 tr s s s rt s à s s r t t r st t rés r é t sté t s é r s st s r é s s r s r t s s s r ss t s s 3 s ér rt s ès s s s r r t rs tr t t s s r s t r t ts 2 rs à s s str t r s r ss t à s t s s s ss t r r r s ts tr s s s rt s s r s ts s é t r s s ét ss t tr s s s rt s t s s tr r s r t s s s s t ts st s r é s r tt r r t é r r ss t r s é t q q é t à s s r r s 2 s ét t à rt r tr r t é sq à r r s s t ts tr s s tr r s à s s P r r r r ss st s r s r s tr r r r s r t s r r ès s s s é ôts è s ét t à rt r s s s rt s t t r t ré t t r s s s s t rt t s s s rt s r t r s 3 s r s s tr s s tr r s s s st rr s st tt t r t s s 1 s r r t s rq s s t s r és r r s t r ré t t r s tr s r s rés t t ê r r t t s r s t ê t t 1 r ss α s rés t ts q t q st t r t s ré rs rs 1 r r s t r s s q s t û à ér ré t
120 t r s é s st q s t été été s r 2s 1 r ss s è s t t s 2 r 12 r q s t r t s ér rs s r q s r P r r 2s s è s tr q s t è t2 q é s s s s s q t r t s s s t è s t2 t r s s s s é ôts r 1 s 2 r st s ré é és r α ss s r st s rt 1 t t t s s ét é s s t rés ts s r r t t r r té st s r é tr s 1 r r rés st t s r r t tr t t ès s s s s t rés ts t r s s s rt s t trés s s t r s s tr t é t r r s s s t r 1 2 s s 2 r st q s 3 s 1 r t t t s 2 r st s st ré t rt t q s r r t s 1 r ts q t 1 q r ér r s é t q s r é 1 r ss β r t r r é q st té s s r s t t s r s r s ér t q s s s s ê s rr t r r 2 r t s t tr q s s r s t β 3 s 1 tr tés r s t s t é s s s s r sé s t ts t s 2 s 2 r st s s r t s q s r s t s 2 r st s q 1 r t s s s rr t q s s 1 r t β st 1 r é r s s ré rs rs s ss r s s 2 r st q s q 1 q t t r s rt t t r s r s s s rt t à r ès s rt t s r P r 1 q r t t été s sé q s s ré rs rs rr t t r tr s t é t é és 2 t s s s s rr t q s s 1 tr tés r s s rq s r s t s r s ttr é s s ré rs rs 1 r t s rq rs és 2 t 1 s ét s tr t été rs s s r s s ré rs rs s é r é r r té tr s ér t 2 r st s sq s r t s s t ttr é s s s s t s ér r rs é t2 é t 2t r r t s t s t r rsq s s s t st é s β rt té st é é s rt s s s r t s t é t2 r st q r é ét t
121 r s α t è t2 1 r ss s α t s è s t t 1 ér rq r é t é t t ré r ss r ér β s tr î r tr s t é t é és 2 t s s s s ér t q s t rs r é q s r ts r s s s s s s ésés rr t tr r èr t r à q s t r s s é t2 r s 2 r st s rt t 1 é ôts è s t à r ès
122 LIVER PRECURSOR CELLS INCREASED HEPATIC FIBROSIS INDUCED BY CHRONIC CARBON TETRACHLORIDE INTOXICATION IN RAT Marie-Noele CHOBERT 1,2, Dominique COUCHIE 2,3, Agnès FOURCOT 1,2, Elie-Serge ZAFRANI 1,2,4, Yannick LAPERCHE 1,2, Philippe MAVIER 2, Arthur BROUILLET 1,2 (1) INSERM, UMR-S-955, Equipe 17, Créteil, France (2) Université Paris-Est Créteil, France (3) INSERM, UMR-S-955, Equipe 21, Créteil, France (4) AP-HP, Groupe Henri Mondor-Albert Chenevier, Département de Pathologie, Créteil, France Corresponding Author: Arthur BROUILLET, INSERM U955 Equipe 17, Institut Mondor de Recherche Biomédicale, Créteil, F-94010, FRANCE Tel : +33 (0) Fax : +33 (0) arthur.brouillet@inserm.fr
123 Introduction Hepatic fibrosis is the major complication of virtually all types of chronic liver damage (including viral, alcohol-related and non-alcoholic fatty liver disease). Even though the primary targets of injury are hepatocytes within the lobule, fibrosis that ultimately leads to cirrhosis initiates predominantly in the portal area in human disease [1]. When the replicative capacity of most remnant hepatocytes is impaired, an alternate regenerative process arises in the portal areas through a ductular cell reaction including liver progenitor cells (LPC). These cells (also called oval cells in rodent) once activated, proliferate and migrate into the hepatic lobule, where they most probably undergo further differentiation into both hepatocytes and bile duct cells to repopulate the hepatic parenchyma. LPC have been of increasing interest in latest years because of their potential role not only in regeneration but also in fibrogenesis and carcinogenesis. In experimental models, LPC expand in close proximity to activated hepatic stellate cells (HSC) producing extracellular matrix [2] and there are increasing data demonstrating an intimate crosstalk between these both types of cells [3-5]. Moreover, studies in rodent models and human diseases have pointed a strong correlation between severity of fibrosis and the intensity of the ductular reaction, suggesting that LPC could play a key role in liver fibrogenesis [6-9]. This correlative relationship between HSC and LPC is not completely understood and their activation could occur independently, successively or in tandem through similar stimuli [10]. First, we could consider that secretion of profibrogenic cytokines and growth factors from LPC could participate to portal fibrosis by inducing HSC proliferation and activation [11, 12], but this direct implication of LPC to fibrosis has not been clearly shown in vivo. Other data underline that activated HSC are a major source of growth factors promoting LPC expansion [13]. Recently, it has been demonstrated that extracellular matrix deposition by myofibroblasts occurred as an initial phase, prior to LPC expansion in the portal region and in front of LPC along the porto-veinous lobular invasion and differentiation [14]. In this case, HSC activation is a pre-requisite for LPC activation, similar to other stem cell niches [15]. Additionally, LPC-HSC cross-talk could enhance fibrogenesis indirectly by recruiting cells of innate immune response such as leukocytes [16]. Among the large variety of factors secreted by HSC and LPC, TGF-β is likely the most potent fibrogenic cytokine. Interestingly, TGF-b is also a wellestablished mediator of epithelial-mesenchymal transition (EMT) which contributes to fibrosis in numerous tissues following injury including the liver [17-21]. Lastly, recent studies illustrate an unexpected plasticity of these both types of cells, adding complexity to the current understanding of fibrogenesis with the possibility that HSC may transdifferentiate into progenitor cells by mesenchymal transition [22] and that LPC display a mixed epithelial/mesenchymal phenotype with characteristics of activated HSC including collagen secretion [23, 24, Wang, 2009 #1053]. From these observations, we hypothesized that the expansion of LPC in portal areas would contribute to fibrogenesis in the liver. To investigate the fibrogenic potential of LPC, we established a new model of fibrosis in the rat where carbon tetrachloride (CCl 4 ) injury was combined to a 2-acetylaminofluorene (AAF) treatment. Chronic
124 CCl 4 injection is a classical model of rodent liver fibrosis [1], initiated by metabolism of CCl 4 into highly reactive CCl 3 radicals by cytochrome P450 in liver parenchymal zone 3 and causing hepatic centrilobular necrosis. In this experimental model, matrix deposition starts around the central with gradual formation of septa bridging central veins without any ductular reaction [25]. In our model, the proliferative hepatocyte response caused by CCl 4 -induced necrosis has been blocked by administration of AAF prior to and during hepatic injury [26] leading to the emergence of an alternate regenerative pathway with expansion of LPC in periportal regions as commonly reported in human fibrosis of various etiologies [6-8, 27]. The involvment of LPC activation in fibrosis was analyzed by comparing the fibrogenic response in rats treated with a combination of CCl 4 and AAF (AC group) to that obtained in animals treated with either CCl 4 (group C) or AAF (group A) independently. We also investigated whether LPC may directly participate to fibrogenesis and serve as a source of myofibroblasts through epithelial-to-mesenchymal transition (EMT) as reported for hepatocytes and biliary cells [17-21]. We also follow the potential of TGFβ to induce an EMT of HepaRG liver progenitor cells in vitro.
125 Materials and Methods: Animals and experimental design. 200g Male Sprague Dawley rats (R. Janvier Animal Center, Le Genest-ST-Isle, France) received intraperitoneal injection of diluted CCl 4 1/1 in olive oil (first injection at 2 ml/kg body weight, next at 1 ml/kg body weight) twice a week in combination with discontinuous 2- acetylaminofluorene (AAF, 0,02% in pellet form) treatment (AAF/CCl4 group, AC). AAF was given five days before the first CCl4 injection and for ten days after. From the second week, the AAFtreatment was discontinued 3 days a week in order to reduce mortality. Rats were sacrificed 2, 4 or 6 weeks after the first CCl4 injection, 48 hours after the last CCl4 injection. Two other animal groups received either CCl4 only (CCl4 group = C) or AAF only (AAF group = A). Six animals were analyzed per time point and per group. An additional group of untreated rats was used as a control and fed a standard rat chow ad libitum (control group, n=7). The livers were removed from animals under anaesthesia and treated as previously described [28]. Blood samples were collected to measure serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities by using an Advia 1650 automate (Bayer Diasys). Animal manipulations were performed according to the recommendations of the French ethical committee and under the supervision of authorized investigators. Histological and immunohistochemistry. Liver injury and collagen content were assessed on 4 µmthick paraffin-embedded liver sections stained with hematoxylin and eosin (H&E) and Sirius red respectively. Liver fibrosis was graded according the METAVIR scoring system. Immunohistochemistry was carried out on paraffin-embedded or 5 µm-thick frozen sections as previously described [28]. Primary antibodies were mouse monoclonal anti-cytokeratin 19 (CK19) (1:150, Novocastra laboratories, Newcastle, UK), anti-alpha-smooth muscle actin (αsma) (1:1000, Sigma-Aldrich, Saint Quentin Fallavier, France), anti-desmin (1:200, DakoCytomotion, Trappes, France), anti-cd68 (1:150, Serotec Cergy Saint Christophe, France) and rabbit polyclonal anti-collagen 1 (1:200, Serotec). Fluorescent labeling of CK19 and CD68 was achieved on frozen sections using secondary goat anti-mouse IgG Alexa fluor 488 and 555 respectively (1:1000, Invitrogen), after an incubation with the Image-iT TM FX signal enhancer (Invitrogen) to reduce the background. For immunodetection of α-sma, sections were revealed by the streptavidin-alkaline phosphatase conjugate (Serotec) and the FastRed substrate system (Dako). Immunofluorescence double staining using co-incubation of rabbit anti-collagen1 and mouse anti-ck19 were revealed respectively with the secondary goat anti-rabbit IgG Alexa fluor 555 and goat anti-mouse IgG Alexa fluor 488. When mouse primary antibodies anti-α-sma or anti-desmine were used, they were incubated first alone and detected with goat anti-mouse IgG Alexa fluor 555. After a 20 minutes step of permeabilization by 0.2% triton in PBS, slides were then incubated with second anti-ck19 antibody directly labeled using Zenon labeling technology (Invitrogen) with mouse IgG Alexa fluor 488.
126 In situ hybridization. The complementary RNA probe specific for AFP and TGFβ were synthesized using a digoxigenin RNA labeling kit (Roche Diagnostics, Meylan, France) from the mouse I.M.A.G.E. cdna clone digested with SalI (antisense probe) or PvuII (sense probe) and transcribed from the T7 or SP6 promoters, respectively. The size of the RNA probes (respectively, 760 bp and 565 bp for the antisense and sense probes), labeling efficiency were checked as previously reported. Hybridization procedures and revelation were performed as previously described [29]. Quantitative real-time PCR. Total RNA was isolated from frozen hepatic tissue using the RNeasy mini kit (Qiagen) and 2 µg were reverse-transcribed from random hexamers using a first-strand synthesis kit (Fermentas, Life Sciences) and specific cdna amplifications were performed as previously described [28] using primers listed in Table 1. Analyses of gene expression were performed with the 2 -ΔΔCT method by using ribosomal 18S RNA normalisation and relative quantification of target mrna was expressed as fold induction over values obtained from control animals. Liver collagen content. Hydroxyprolin assay was performed as described [30] using small fragments of different lobes that were pooled, lyophilized, and hydrolyzed in 6 N HCl at 110 C overnight and hepatic hydroxyprolin content was spectrophotometrically measured by reacting with freshly prepared Ehrlich s reagent and results are expressed in micrograms per milligram of tissue (dry weight). HepaRG Cell Culture. HepaRG cells were cultured as described [31]. Cells ( /cm2) were maintained in William's E medium supplemented with 10% fetal bovine serum, 100 U/ml penicillin, 100 g/ml streptomycin, 5 g/ml insulin, and M hydrocortisone hemisuccinate. To obtain HepaRG differentiation, cells were cultured in the medium for 2 weeks and EMT was induced by addition of TGFb for 2 more weeks. Statistical analysis : All data are expressed as mean ± SEM. Two-way ANOVA was employed to assess the effect of each treatment over the weeks and the non-parametric Mann-Whitney U-test was used to compare the differences between treatments in each time period. Statistical analyses were performed with PRISM (GraphPad) and P-value of less than or equal to 0.05 indicated a significant difference between groups.
127 Results Liver injury CCl 4 treatment increased gradually serum ALT and AST activities (Figure 1A) which reached maximal values at 6 week (238±33 and 328±27 U/L respectively). No significant differences were observed between the C and AC animals by two-way ANOVA illustrating similar liver injury in both groups, even if ALT was significantly higher in group C at the later stage (6 weeks). Interestingly, animals treated with AAF alone had significantly elevated ALT and AST values (52± 8 and 112±6 U/L respectively; P< 0.05) when compared with control rat (37±5 and 90±5 U/L) revealing that administration of AAF alone also induces a moderate hepatocellular necrosis. In a preliminary study, a continuous AAF exposition in association with usually CCl4 dose at 2ml/kg of body weight were not well tolerated leading to high rate of animal mortality and severe weight loss after 4 weeks of treatment (data not shown). For these reasons, after the first injection of CCl4 at 2ml/kg of body weight, we performed the subsequent injections with half dose of CCl4 (i.e. 1ml/kg) and the AAF pellets were replaced by standard chow 3 days per week. Livers from CCl 4 -treated animals exhibited gradual hepatic parenchymal alterations in centrilobular areas that correlated to the length of the treatment (figure 1B), in accordance with increased serum levels of transaminases. After 2 weeks of CCl 4 treatment, focal necrosis with vacuolated hepatocytes and inflammatory cells infiltration was seen around the central venules (data not shown). Two weeks later, livers showed mild to moderate steatosis often associated with inflammatory cell accumulation and bands of damaged tissue connecting central vein of the adjacent lobule (Figure 1B). After 6 weeks, the hepatic lobular structure was disorganized with marked steatosis and ballooning degeneration of hepatocytes in close proximity of infiltrated cells, leading in some cases to complete interconnection between central veins and thus, dividing the parenchyma into pseudo-lobules centered on a portal tract. In liver sections from the AC group after 2 weeks of treatment, hepatic centrilobular injury was similar to that observed in animals treated with CCl 4 alone, but additionally, an expansion of basophilic cells including cells with small ovoid nuclei (oval cells), not shown in the C group, was detected in periportal areas (data not shown). After 4 weeks, this ductular reaction extended more deeply into the lobule and formed portoportal bridges and, two weeks later, large bands of infiltrated basophilic cells delineated multiple nodules in the liver (Figure 1B). Moreover, steatotic hepatocytes were localized around these connecting septa and in comparison with group C, this macrovesicular steatosis was more severe at 4 weeks of treatment. Unexpectedly in the group A, we observed a proliferation of basophilic cells in periportal areas progressing around the lobule and forming porto-portal bridges after 6 weeks of treatment, although liver parenchymal architecture was almost preserved with neither steatosis nor necrosis in centrilobular areas in contrast with the two other groups. Liver precursor cell expansion
128 In groups A and AC after 2 weeks of treatment, small basophilic cells with a high nuclear/cytoplasmic ratio arising from portal areas were stained with cytokeratin 19 (CK19), a liver precursor cell (LPC) marker (data not shown). In these two groups, CK19-positive LPC increased in number with time and expanded inside the lobule where they were organized in small clusters of three to four cells (Figure 2A). At 6 weeks in both groups, there was an extensive arborization of expanding duct-like structures, which extended across the liver lobule, forming bridges between portal tracts. In addition in group AC, marked LPC expansion produced connections between portal and centrilobular areas, dividing the parenchyma into smaller pseudo-lobules. As expected, such an accumulation of CK19-positive LPC was not detected in CCl 4 -treated rats, where labeling was restricted to bile duct-cells in portal tracts. These results were confirmed by real-time PCR analysis revealing an increase of hepatic CK19 mrna expression only in A and AC groups, peaking at 84-fold over the basal level at week 4 and 6 in AC-treated rats (Figure 2B). No significant difference was observed between the A and AC groups, revealing that CCl 4 treatment did not take part to the magnitude of LPC response induced by AAF. In line with this result, AFP mrna was localized by in situ hybridization in cells of the ductular reaction, convincingly confirming their precursor phenotype (Figure 2C). In summary, chronic AAF treatment activated the hepatic stem cell compartment and the accumulation of LPC was similar in groups A and AC. Differences in the parenchymal localization of LPC, notably in porto-central connections in the group AC seemed related to CCl 4 -induced architectural distortion. Liver Fibrosis As expected, rats treated with chronic CCl 4 injection displayed progressive liver fibrosis (Figure 3A) detected by sirius red staining (revealing accumulated collagen, predominantly type 1 & 3). At week 2, mild fibrosis started and extended from hepatic central venules with deposition of thin collagen fibers in necrotic and steatotic areas. Later at week 4, we observed gradual formation of fibrous septa interconnecting two centrilobular areas without disorganization of lobular architectur. After 6 weeks, multiple centro-central fibrotic septa incompletely compassed and divided hepatic lobules. In animals treated by combination of AAF and CCl 4, the disease progression and the fibrotic pattern were completely different. In addition to centrilobular fibrosis, also observed in CCl 4 -treated animals at 2 weeks, sirius red-staining revealed collagen deposition extending from portal fields and surrounding the ductular reaction in group AC. Two weeks later, some thin fibrous septa from the portal tracts were interconnected with larger fibrous scars from central veins, leading to porto-central bridging fibrosis (METAVIR F2-F3). After 6 weeks, fibrosis was more extended and delineated multiple micronodules leading up to complete cirrhosis (METAVIR F4) in all rats from AC group. These data indicate that AAF, by stimulating an expansion of LPC, exacerbates CCl 4 -induced liver fibrosis. In support of this result, hepatic mrna expression of collagen type 1 and 3 was largely and significantly increased in the group AC in
129 comparison with CCl 4 -treated animals (ANOVA p<0.05). This more pronounced fibrosis was also confirmed by an increase of the hydroxyprolin content (Figure 3C) in AC group (up to 7 fold over basal level at week 6, ANOVA p<0.001) in accordance with a higher METAVIR score. Unexpectedly in AAFtreated animals after 2 and 4 weeks, thin bands of collagen were detected arising from portal tracts and progressing around the lobule at week 6 in close proximity to the LPC expansion (Figure 2A), suggesting that this faded signal may correspond to type 4 collagen [32], a major component of basement membrane associated to LPC activation and migration within the hepatic lobule [33]. In accordance with this observation, real-time PCR analysis revealed an induction of hepatic Col4 mrna which paralleled the number of CK19-positive cells in A and AC groups (Figure 3B) and which was not observed in CCl 4 - treated animals. In A group, livers exhibited a minor induction of collagen 1 & 3 mrna expression which was significantly lower than in CCl 4 -treated animals, confirming that sirius red staining in the AAF group corresponds mainly to type 4 collagen. Thus, the 2-times induction of the hydroxyprolin content in the AAF group may represent the extension of basement membrane associated with LPC expansion. Moreover in AAF-treated animals, laminin, another component of the basement membrane, surrounded LPC in periportal areas, all along their spreading into the parenchyma (group A) and in perinodular areas of AAF/CCl 4 -treated animals (data not shown). Therefore, these results support active deposition of basement membrane components during LPC expansion, could also take part in the process of fibrosis in AC group. Liver inflammation Liver injury induced by CCl 4 injection generates an inflammatory response and macrophages play a critical role into the myofibroblastic transformation of HSC and consequently in hepatic fibrogenesis [1]. The amount of liver macrophages, including Kupffer cells and recruited monocytes, was assessed by detecting the pan-macrophage CD68 antigen. The normal liver showed a homogenous lobular distribution of CD68-positive cells in accordance with expected Kupffer cells localization within sinusoids and a computer-assisted spot detection demonstrated that CD68 staining represented 2.1 ± 0.5 % of the liver section (data not shown). After 4 and 6 weeks of CCl 4 injection, aggregates of CD68-positive cells accumulated in centro-central fibrotic septa (Figure 4A) and the staining increased with the duration of treatment and covered up to 6.3% of the liver section (Figure 4B). In animals treated with only AAF exhibiting no fibrosis, we observed an induction of CD68-labelling areas which was not significantly different to C group (Figure 4B), suggesting that macrophages may not be the only cells responsible for hepatic fibrogenesis. Furthermore in A group, most positive cells were detected all over liver parenchyma, identically to the CD68-stained Kupffer cells in control sections of normal liver and some macrophages were accumulated close to LPC expansion in portal areas (Figure 4A). In animals treated by combination of AAF and CCl 4, a majority of CD68-positive cells was accumulated around perinodular
130 areas (Figure 4A), in the middle and margin of the bridging septa divided cirrhotic liver described above (Figure 2A). Although extensive fibrosis was detected in AC group, the percentage of the liver tissue that stained for CD68 did not significantly differ from C group (Figure 4B). During liver injury, activated macrophages secrete numerous cytokines, including TNFα which are mitogenic for hepatocytes [34] as well as for LPC [35, 36]. In CCl 4 -treated rats, RNA quantification by real-time PCR revealed a gradual but limited (less than 4 fold) induction of TNFα mrna (Figure 4C) to a level which was not statistically different from A and AC groups. Even more, CCR2 mrna expression which is limited to recruited monocytes and some T lymphocytes [37], was not different in the 3 groups of animals whatever the treatment (Figure 4D). Taken together, these results revealed a modest inflammation in liver after a CCl 4 and/or AAF treatments. The low and comparable grade of inflammation in the 3 groups, is in contrast with the strong difference in fibrosis suggesting that, in our experimental conditions, inflammation was not a major player in the fibrogenic process. Fibrogenic liver cells During chronic liver pathologies, portal fibroblasts and activated HSC exhibited phenotypic features of myofibroblasts, namely expression of αsma and extracellular matrix (ECM) proteins, [1]. After 2 and 4 weeks of CCl 4 treatment, immunohistochemical detection of αsma revealed only a perivascular labelling (Figure 5A), also detected in control livers (data not shown). At week 6, αsma-positive cells were observed in active fibrotic areas (Figure 5A B) in only 3 treated-animals out of 6 analyzed. In contrast, in the AAF/CCl4 treated-group, numerous αsma-positive cells were seen around portal areas and infiltrated in the lobule as early as after 2 weeks of treatment (Figure 5A and 5B). Later, a strong αsma labelling was also revealed in fibrous septa (at week 4) and around perinodular areas (at week 6) in liver of AC-treated animals with excessive ECM deposition leading to cirrhosis. In the AAF group, whatever the time of treatment, the αsma-staining was restricted around the vessels as in normal liver and did not reveal any activated HSC in ductular reaction (Figure 5A). Immunofluorescence detection of αsma in liver frozen sections (which provides a higher sensitivity as compared to immunodetection on paraffin sections) confirms the lack of αsma-positive cells in the ductular reaction from AAF-treated animals (Figure 6A) and the presence of few activated HSC in fibrous septa in liver from C group. Immunofluorescence localization of desmin, a hallmark of quiescent and activated HSC showed an accumulation of positive cells in the AAF group, closely associated to LPC expansion and extending from periportal areas (Figure 6A). In the CCl4 group, desmin staining was detected in centrilobular injured areas and in centro-central fibrous septa. Thus, under our experimental conditions of CCl 4 injection, the paucity of αsma staining reflected minor myofibroblastic activation, in agreement with moderate fibrosis. Nevertheless, in the C group where HSC (Desmin +, αsma +/- ) were not fully activated, we observed an accumulation of collagen 1. (Figure 6A). In cirrhotic liver of AC group where HSC are fully
131 activated, we observed large amount of collagen 1 deposition with a fluorescent signal distribution similar to αsma labelling in perinodular areas. In A group where HSC (Desmin +, αsma - ) are not activated, a faint collagen 1 deposition was observed around the ductular reaction. In summary, HSC transactivation was only evidenced in liver of AC group, first in close proximity of LPC arising from portal areas and then in connecting septa, demonstrating that combination of CCl 4 with AAF was necessary to obtained αsma-positive myofibroblasts which could account for high level of fibrosis in this group. To identify more precisely the source of collagen I around the LPC, we performed double immunofluorescent staining for CK19 and collagen 1 in cirrhotic liver from AC group after 6 weeks of treatment (Figure 6B). CK19-positive LPC, whether organized in duct-like structures around portal areas or in cluster in fibrous septa, were surrounded by collagen I fibers. No colocalization was observed between the cytoplasmic marker of LPC and this secreted matrix component, suggesting that the primary cells responsible for liver fibrogenesis are HSC surrounding the LPC. Among many cytokines involved in liver fibrosis, TGFβ produced by activated HSC appears to be the most important one [1] and interestingly, its expression coincides with the LPC expansion during liver regeneration after AAF/partial hepatectomy in rats [11]. Real-time PCR analysis demonstrated that hepatic mrna expression of this profibrogenic factor was significantly higher in AAF/CCl 4 group (p< 0.05) than in CCl 4 alone (Figure 6C), in accordance with the evaluated fibrosis score. Unexpectedly, induction of TGFβ mrna was also more pronounced in group A than in C (p< 0.001), notably after 2 and 4 weeks of treatment corresponding to early stages of LPC activation. An increased TGFβ expression in both groups treated with AAF suggests that LPC are themselves a source of TGFβ, a result which have already been evocated [38, 39]. To identify more precisely the TGFβ-producing cells, we performed in situ hybridization studies (Figure 4D). As expected in the liver of animals treated by CCl 4 during 6 weeks, a small number of TGFβ-expressing cells accumulated in fibrous septa beside steatosic alterations (Figure 5D) in areas where we have detected few αsma-positive myofibroblastic cells (Figure 5A). In contrast, in rats treated with AAF alone during 6 weeks, numerous cells arranged as duct-like structures that deeply invaded the liver from portal areas were positive for the expression of TGFβ mrna (Figure 5D). In this group without αsma-positive myofibroblastic HSC (Figure 5A), TGFβ expression takes place in cells previously identified as LPC by CK19 staining (Figure 2A) and AFP expression (Figure 2C). In cirrhotic liver of AAF/CCl4-treated animals, TGFβ mrna expression was detected in perinodular septa, in cells almost always organized in duct-like structures characteristic of the ductular reaction (Figure 6D) but a part of the signal could also be attributed to αsma-positive myofibroblasts closely associated to LPC expansion (Figure 5A and 6A). These results support real-time PCR analysis demonstrating induction of TGFβ mrna expression (Figure 6C) in correlation with number of activated HSC and/or LPC that emerge in liver after each treatment. Thus, the 5-fold induction of TGFβ mrna after 6 weeks of treatment was related to an expression by HSC in animals treated by CCl 4 and by LPC in
132 animals treated by AAF alone. The further induction up to 12 times in AC group was related to TGFβ expression by both LPC and HSC as a consequence of AAF-induced LPC expansion and CCl 4 -induced myofibroblastic activation. In addition, the most potent fibrogenic cytokine TGFβ produced by LPC in the AC group can induce the production of ECM by the neighbouring activated HSC. From these observations, we conclude that LPC contribute to the development of liver fibrosis. LPC do not express mesenchymal markers To determine whether LPC could undergo an epithelial-to-mesenchymal transition (EMT) in cirrhotic liver, we analyzed these CK19-positive cells for the expression of αsma, desmin and FSP1 in AC group after 6 weeks of treatment. Double immunofluorescent experiments did not evidence expression of these mesenchymal markers in CK19-positive LPC either in the ductular reaction arising in portal areas or in fibrous septa (Figure 7A). Then, it can propose that αsma-positive signal found in close contact of LPC correspond to the labelling of myofibroblastic HSC already described in the ductular reaction [2, 9, 14, 40]. Desmin staining also revealed HSC in front of LPC, but it extended beyond αsma-labelled cells (Figure 7A), indicating that several HSC detected in the sinusoidal space far of the ductular reaction are not fully activated into the myofibroblastic phenotype. These results were confirmed by a confocal analysis which clearly showed the absence of colocalization of αsma or desmin in CK19-labelled cells (Figure 7B). Additionally, expression of the fibroblast specific protein FSP1 which was also revealed around CK19-positive LPC (Figure 7A) did not co-stain in same cells. Therefore, we did not obtain any evidence of LPC undergoing EMT in injured liver from AAF/CCl 4 -treated rats. TGFβ-induced EMT in HepaRG cell line. In our in vivo study, we could not highlight expression of mesenchymal markers in LPC, but transition of epithelial LPC from a duct-forming structure into isolated myofibroblasts could be difficult to identify in injured liver. Moreover, we could not exclude the possibility of confounding these cells acquiring expression of mesenchymal components, with activated HSC which are closely associated with LPC expansion. To further address the issue of EMT, we cultured HepaRG liver progenitor cells and followed in vitro their epithelial and fibroblastic phenotypic characteristics in response to TGFβ. HepaRG is a recently described cell line obtained from liver tumor tissue of a patient suffering from hepatitis C virus associated hepatocellular carcinoma [31, 41]. These cells express specific hepatocyte function, displaying both hepatocyte-like and biliary-like epithelial phenotypes and expressing LPC specific markers. Seven days after plating, HepaRG cells showed a typical epithelial morphology displaying a polygonal shape while confluence was reached (Figure 8A). When cells were stimulated with TGFβ (X ng/ml) for 7 days, the majority of cells that survived acquired a fibroblastic phenotype characterized by a more elongated
133 form (Figure 8B). Both αsma (Figure 8D) and collagen I (Figure 8F) were strongly expressed in TGFβtreated cells whereas staining of these mesenchymal markers was very faint in unstimulated cells (Figure 8C and 8E). To characterize more precisely this TGFβ-induced EMT of HepaRG revealed by changes in morphology and cytoskeleton rearrangement, we quantified mrna coding for both epithelial and mesenchymal markers (Figure 8G). We found that collagen 1 & 3 expression was dramatically induced in response to TGFβ treatment. In accordance with immunofluorescent analysis, a higher level of αsma mrna was detected in TGFβ-stimulated cells as compared to their controls. In contrast and as expected during EMT, expression of E-cadherin, a universal epithelial marker that plays a key role in the maintenance of cell junctions, was largely downregulated after TGFβ treatment. Moreover, an induction of Snail, a repressor of E-cadherin transcription, was also consistent with EMT. In conclusion, TGFβ is able to induce EMT of HepaRG, suggesting that this fibrogenic factor produced by LPC during liver injury could contribute to the fibroblastic transformation of these cells which could directly participate to collagen deposition and hepatic fibrogenesis.
134 Discussion It is now well documented that in human liver, progenitor cell compartment is activated in all chronic liver diseases and that severity of fibrosis correlates with LPC expansion [6-9, 27]. In this study, in order to evaluate the importance of LPC in hepatic fibrosis, we developed a new experimental model of chronic liver diseases with LPC expansion. In rodent models, activation of the LPC compartment is observed when proliferative capacity of hepatocyte is suppressed by the injuring agents and in rat, AAF treatment combined with partial hepatectomy (AAF/PH) or with unique CCl4 injection are among the most widely experimental models used [26, 33]. In both models, successful repair involves a regenerative process in which LPC may contribute to a complete and normal liver restoration after about 2 weeks. Models of injury based on PH or single CCl4 injection lead to a regenerative process without LPC expansion and are therefore not suitable to investigate the effects of LPC on fibrogenesis. Hepatic fibrosis is the result of persistent liver injury and incomplete resolution of a normal wound healing process. Thus, we combined an AAF treatment to repeated CCl4 injections, a well-characterized model of hepatic fibrosis. A difference in tolerance of the chronic treatment was observed between the 3 groups as revealed by the weight of animals after 6 weeks of treatment from AC group (262 ±20 g) which was significantly lower than in groups C and A (370±28 g and 367±16 g respectively). Unexpectedly, chronic AAF treatment by itself induces a true LPC expansion in absence of an external mitogenic stimulus. Yet impaired hepatocyte proliferation during liver growth in young 7-week old rats might provide a sufficient signal to stimulate the LPC compartment. Moreover, a toxicity of AAF could not be excluded during chronic AAF treatment as revealed by slight induction of ALAT/ASAT over 6-week treatment period in spite of undetectable hepatic necrosis. These findings suggest that AAF-treated animals were able to compensate a lack of hepatocyte proliferation by the induction of an alternative regenerative process involving LPC to finally reach a similar liver mass than in the CCl4 group. In the AC group, after 6 weeks of treatment,, we observed a worsened fibrosis as compared to the C group revealing that LPC-mediated regeneration exacerbates the progression of liver fibrosis. An accumulation of LPC in fibrous septa was associated with enhanced activation of HSC and expression of profibrotic genes such as Col1&3, leading to excessive ECM deposition with established cirrhosis in AAF/CCl4-treated animals. Despite similar liver injury in both groups, chronic CCl4 treatment alone is not sufficient to promote LPC expansion. Interestingly, LPC expansion which occurred in rats treated with AAF alone, was not accompanied with αsma-positive HSC and excessive ECM deposition, except an enhanced expression of Col4 and accumulation of laminin (data not shown), two components of the basement membrane in contact with LPC [2]. These data suggest that LPC alone are not fibrogenic but emphasize the need of CCl4-induced injury to promote HSC activation and subsequent fibrosis. Indeed, HSC can be activated by the engulfment of necroapoptotic hepatocytes [42]. Recently, it was demonstrated that attenuated hepatocyte proliferation in Egr1 -/- mice after chronic CCl4 exposure drives
135 the activation of LPC [43] which contributes to enhanced hepatic fibrosis and confirms the role of LPC activation in the fibrogenic process. LPC-mediated liver regeneration is a complex process, involving cytokine secretion and remodelling of the extracellular matrix [13]. Many experimental models of LPC-mediated liver regeneration have reported co-activation of LPC and HSC and recent data clearly established that HSC activation is required for LPC response. During a choline-deficient ethionine supplemented diet, αsma-positive cells and collagen deposition occur at day 3, prior to LPC expansion, revealing that activation of HSC arranges an hepatic microenvironment favorable to LPC proliferation and differentiation [14]. Interestingly in the AAF/partial hepatectomy model, it has been shown that inhibition of HSC activation either by iloprost or L-cysteine greatly diminished LPC expansion and their differentiation into hepatocytes, most likely by the lack of an appropriate microenvironment within the regenerating zone [44]. HSC are a major source of growth factors for LPC, but inflammatory cells such as Kupffer cells are likely to provide additional cytokines mitogenic for LPC, including TNFα [12, 45]. Resident and recruited macrophages play a critical role into the myofibroblastic activation of HSC and consequently in hepatic fibrogenesis notably by releasing TGFβ [1]. In our study, the similar amount of liver macrophages, as well as the unchanged hepatic level of cytokines such as TNFα, could not account for the difference of intensity of LPC response and hepatic fibrogenesis observed between the 3 experimental groups. Clearly, the fibrogenic process could not entirely depend on inflammatory response which was moderate in our experimental setting. Numerous studies have revealed the close proximity of activated HSC to LPC but it does not prove that LPC directly cause HSC activation and fibrosis. Other interpretation could be that stimulation of LPC and HSC occurs independently, perhaps through similar mediators or stimuli [10]. While our study does not resolve this question, we demonstrated in AAF/CCl4-treated rats that LPC express the most potent fibrogenic growth factor TGFβ, an activator of quiescent HSC into myofibroblasts [1]. Accordingly in these animals, we detected many αsma-positive HSC closely associated with LPC expansion in relation with excessive liver fibrosis. Thus, LPC, by expressing TGFβ, could directly participate to the myofibroblastic activation of HSC and to subsequent fibrosis in injured liver from AAF/CCl4-treated rats. Although TGFβ was also detected in LPC from rats treated with AAF alone, we could not evidence αsma-positive cells neighbouring LPC suggesting that TGFβ alone is not sufficient to induce activation of HSC when there is no damage of the hepatocytes. The role of EMT in fibrosis was established in many organs including the liver and TGFβ-induced EMT has been demonstrated with hepatocytes, biliary cells and HSC [17-21]. The plasticity of HSC that coexpress mesenchymal and LPC markers [46] and their putative differentiation into hepatocytes through mesenchymal-epithelial transition [22], add complexity to the contribution of HSC and LPC to liver fibrosis. However, the ability of LPC to provide a source of myofibroblasts through EMT [46, Yang, 2008 #990, Wang, 2009 #1053] is yet uncertain [47]. We could not evidence LPC undergoing EMT in vivo in injured liver since we did not localize expression of mesenchymal proteins in CK19-positive cells.
136 However, CK19-negative cells expressing mesenchymal markers are detected in close proximity of LPC indicating that the primary cells responsible for liver fibrogenesis in AAF/CCl4-treated animals are HSC and that LPC may contribute to their activation by secreting the profibrogenic factor TGFβ. We did not observe any evidence for an EMT of LPC in vivo. A rapid loss of CK19 expression during the EMT of non-polarized cells could render the identification of transitioning LPC difficult in vivo. Transition of LPC to myofibroblasts was obtained in vitro using human HepaRG cell line under TGFβ stimulation. these cells underwent a TGFβ-induced phenotypic change from epithelial to mesenchymallike cells with an increased expression of snail, αsma and collagens and a paralleled decrease of E- cadherin. Similar results were obtained with LPC isolated from choline-deficiency ethionine-treated rats, which differentiate into mesenchymal cells through an EMT process induced by TGFβ in vitro [24]. Although isolated and cultured LPC possess some mesenchymal markers ofhsc such as collagen 1&3 [24] and αsma [23, 46], an elegant fate-mapping study using the LPC marker Foxl1 demonstrated in vivo that all Foxl1-expressing cells and their progeny did not acquired a mesenchymal phenotype during bile duct ligation or treatment with DDC promoting LPC expansion [47]. Interestingly in this study, fibroblastic cells (expressing elastin, desmin and αsma) are located in close proximity to Foxl1-positive cells revealing a potential crosstalk between LPC and these fibrogenic cells. In accordance with our in vivo study, these data demonstrate that LPC do not acquire a mesenchymal phenotype through EMT nor directly participate to excessive ECM deposition. Altogether, our study revealed that fibrosis in response to chronic CCl4 intoxication was worsened by LPC expansion induced by AAF treatment. The strong fibrogenic response in AAF/CCl4-treated animals as compared to CCl4 tretment, was not the consequence of altered liver injury or inflammatory response but due to the accumulation of LPC which contributes to fibrosis mainly through the production of TGFβ an inducer of HSC myofibroblastic transformation the major fibrogenic cell type involved in fibrogenesis. Figure legends Figure 1 : A) ALAT and ASAT activities in serum gradually increased after CCl4 administration during 6 weeks and no significant difference was observed in AAF/CCl4-treated rats. Administration of AAF induces a slight hepatocellular necrosis. B) Representative micrographs of the H&E staining following 4 and 6 weeks of treatment (original magnification x40 and x200 as indicated). Figure 2 : A) Immunofluorescent detection of CK19-positive cells in the liver section reveled an accumulation of LPC in rats treated with AAF and AAF/CCl4, arising from portal areas at 4 weeks and expanding inside the lobule after 6 weeks of treatment (original magnification x40, inset shows enlarged images at x200 to better illustrated CK19 labeling). Such an accumulation of LPC was not detected in CCl4 group were CK19 labeling was restricted to bile duct cells. B) Real-time PCR revealed an increase
137 of CK19 mrna expression only in both AAF and AAF/CCl4 groups. C) AFP mrna detected by in situ hybridization (AFP AS probe) was restricted in cells of the ductular reaction in AAF and AAF/CCl4- treated animals. No specific signal was observed using AFP sense probe (original magnification x100). Figure 3 : A) Picrosirius red staining visualizing collagen accumulation revealed progressive liver fibrosis induced by CCl4, with formation of C-C fibrotic septa at 4 weeks and multiple C-C fibrous septa incompletely divided hepatic lobule at 6 weeks. A marked fibrosis with bridging C-P septa at weeks 4, leading up to complete cirrhosis were observed at 6 weeks in AAF/CCl4-treated rats. In portal areas of animals of AAF group, a faint picrosirius red staining was observed around the LPC expansion. Original magnification x100. B) Quantitative RTPCR analysis showed a gradual induction of collagens type 1 and 3 mrna involved in liver fibrosis, which was enhanced in AAF/CCl4-treated rats as compared to CCl4 treatment alone. Induction of collagen type 4 mrna, a major component of basement membrane, was only evidenced in both AAF and AAF/CCl4 groups with LPC expansion. C) Total liver collagen content determined by hydroxyprolin assay was higher in AAF/CCl4-treated rats and correlated with excessive fibrosis revealed either by sirius red staining and collagen mrna expression. Figure 4 : A) Immunofluorescent detection and B) quantification of CD68-positive cells in liver from animals did not reveal significant difference in number of hepatic macrophages between 3 groups following 4 and 6 weeks of treatment (original magnification x100), except a localization all over liver parenchyma in group A and an accumulation of CD68-positive cells in fibrous septa after CCl4 treatment and in the bridging septa divided cirrhotic liver of AAF/CCl4-treated animals. Quantitative RT-PCR did not evidence difference on B) TNFα or C) CCR2 mrna expression revealing that inflammatory response was comparable in the 3 groups. Figure 5 : A) Immunohistochemical detection of αsma-positive cells revealed few activated HSC after CCl4 administration, whereas a strong staining was detected in fibrous septa at 4 weeks and perinodular areas at 6 weeks from liver of. No αsma-positive cells were observed in ductular reaction in AAF group, except the signal from vessels in portal areas. Original magnification x100. B) Higher magnification (x400) clearly evidenced collagen deposition in fibrous septa after 6 weeks of CCl4 treatment, whereas few activated HSC was obtained. In contrast, activation of HSC was detected at 2 weeks in AAF/CCl4- treated rats, around the LPC expansion arising from periportal areas and in bridging fibrous septa at 6 weeks containing large amount of collagen deposition revealed by picrosirius red staining. Figure 6 : A) Immunofluorescent detection of desmin, αsma and collagen 1 at 6 weeks of treatment, confirms the strong ECM deposition in perinodular areas associated with activation of HSC into their αsma-myofibroblastic phenotype in cirrhotic liver of AAF/CCl4-treated rats after 6 weeks. A lower
138 collagen accumulation in relation with very few αsma-positive HSC was evidenced in fibrous septa after CCl4 administration. Lack of activated HSC with faint collagen 1 labelling was observed around the portal areas in the AAF-induced ductular reaction. Original magnification x100. B) No colocalization (with yellow merged signal) was observed with the double immunofluorescent staining for CK19 (green) and collagen 1 (red) in cirrhotic liver from AC group after 6 weeks of treatment (Original magnification x400). C) Quantitative RT-PCR analysis revealed an enhanced induction of TGFβ mrna expression in AAF and AAF/CCl4-treated animals and D) in situ hybridization TGFβ mrna detection revealed a strong positive signal in LPC organized in duct-like structures. Figure 7: Immunolocalization of mesenchymal protein on frozen sections. A) Double immunofluorescent labelling of CK19 and myofibroblastic markers αsma, desmin and FSP1 in AC group after 6 weeks of treatment (x200). Merging signals did not reveal any expression of these mesenchymal markers in CK19 positive cells. B) Confocal analysis evidenced that CK19-positive LPC are surrounded by desmin and αsma positive cells (X400). Figure 8 : Effect of TGFβ on the epithelio-mesenchymal transition of HepaRG cells. Cells were cultured for 2 weeks as described in Method section to obtain their differentiation and for 2 more weeks in absence (A, C, E) or presence of TGFβ (B, D, F) to induce EMT. Cell morphology was observed under phase contrast microscopy (A, B) (X100) and immunofluorecent staining of αsma (C, D) and desmin (E, F) (X200) revealed TGFβ-induced EMT. The level of Col1, αsma, Col3, Snail and E-cadherin mrna (G) assessed by quantitative RT-PCR was expressed as fold induction over values obtained from undifferentiated cells.
139 Bibliography 1. Ramadori G, Saile B. Portal tract fibrogenesis in the liver. Lab Invest 2004;84: Paku S, Nagy P, Kopper L, Thorgeirsson SS. 2-acetylaminofluorene dose-dependent differentiation of rat oval cells into hepatocytes: confocal and electron microscopic studies. Hepatology 2004;39: Friedman SL. Mechanisms of hepatic fibrogenesis. Gastroenterology 2008;134: Fausto N, Campbell JS, Riehle KJ. Liver regeneration. Hepatology 2006;43:S Svegliati-Baroni G, De Minicis S, Marzioni M. Hepatic fibrogenesis in response to chronic liver injury: novel insights on the role of cell-to-cell interaction and transition. Liver Int 2008;28: Lowes KN, Brennan BA, Yeoh GC, Olynyk JK. Oval cell numbers in human chronic liver diseases are directly related to disease severity. Am J Pathol 1999;154: Clouston AD, Powell EE, Walsh MJ, Richardson MM, Demetris AJ, Jonsson JR. Fibrosis correlates with a ductular reaction in hepatitis C: roles of impaired replication, progenitor cells and steatosis. Hepatology 2005;41: Richardson MM, Jonsson JR, Powell EE, Brunt EM, Neuschwander-Tetri BA, Bhathal PS, et al. Progressive fibrosis in nonalcoholic steatohepatitis: association with altered regeneration and a ductular reaction. Gastroenterology 2007;133: Knight B, Akhurst B, Matthews VB, Ruddell RG, Ramm GA, Abraham LJ, et al. Attenuated liver progenitor (oval) cell and fibrogenic responses to the choline deficient, ethionine supplemented diet in the BALB/c inbred strain of mice. J Hepatol 2007;46: Clouston AD, Jonsson JR, Powell EE. Hepatic progenitor cell-mediated regeneration and fibrosis: chicken or egg? Hepatology 2009;49: Pi L, Oh SH, Shupe T, Petersen BE. Role of connective tissue growth factor in oval cell response during liver regeneration after 2-AAF/PHx in rats. Gastroenterology 2005;128: Akhurst B, Matthews V, Husk K, Smyth MJ, Abraham LJ, Yeoh GC. Differential lymphotoxinbeta and interferon gamma signaling during mouse liver regeneration induced by chronic and acute injury. Hepatology 2005;41: Bird TG, Lorenzini S, Forbes SJ. Activation of stem cells in hepatic diseases. Cell Tissue Res 2008;331: Van Hul NK, Abarca-Quinones J, Sempoux C, Horsmans Y, Leclercq IA. Relation between liver progenitor cell expansion and extracellular matrix deposition in a CDE-induced murine model of chronic liver injury. Hepatology 2009;49: Kuwahara R, Kofman AV, Landis CS, Swenson ES, Barendswaard E, Theise ND. The hepatic stem cell niche: identification by label-retaining cell assay. Hepatology 2008;47: Ruddell RG, Knight B, Tirnitz-Parker JE, Akhurst B, Summerville L, Subramaniam VN, et al. Lymphotoxin-beta receptor signaling regulates hepatic stellate cell function and wound healing in a murine model of chronic liver injury. Hepatology 2009;49: Kaimori A, Potter J, Kaimori JY, Wang C, Mezey E, Koteish A. Transforming growth factorbeta1 induces an epithelial-to-mesenchymal transition state in mouse hepatocytes in vitro. J Biol Chem 2007;282: Zeisberg M, Yang C, Martino M, Duncan MB, Rieder F, Tanjore H, et al. Fibroblasts derive from hepatocytes in liver fibrosis via epithelial to mesenchymal transition. J Biol Chem 2007;282: Robertson H, Kirby JA, Yip WW, Jones DE, Burt AD. Biliary epithelial-mesenchymal transition in posttransplantation recurrence of primary biliary cirrhosis. Hepatology 2007;45: Rygiel KA, Robertson H, Marshall HL, Pekalski M, Zhao L, Booth TA, et al. Epithelialmesenchymal transition contributes to portal tract fibrogenesis during human chronic liver disease. Lab Invest 2008;88: Battaglia S, Benzoubir N, Nobilet S, Charneau P, Samuel D, Zignego AL, et al. Liver cancerderived hepatitis C virus core proteins shift TGF-Beta responses from tumor suppression to epithelialmesenchymal transition. PLoS ONE 2009;4:e4355.
140 22. Yang L, Jung Y, Omenetti A, Witek RP, Choi S, Vandongen HM, et al. Fate-mapping evidence that hepatic stellate cells are epithelial progenitors in adult mouse livers. Stem Cells 2008;26: Yovchev MI, Grozdanov PN, Zhou H, Racherla H, Guha C, Dabeva MD. Identification of adult hepatic progenitor cells capable of repopulating injured rat liver. Hepatology 2008;47: Wang P, Liu T, Cong M, Wu X, Bai Y, Yin C, et al. Expression of extracellular matrix genes in cultured hepatic oval cells: an origin of hepatic stellate cells through transforming growth factor beta? Liver Int 2009;29: Zhao XY, Wang BE, Li XM, Wang TL. Newly proposed fibrosis staging criterion for assessing carbon tetrachloride- and albumin complex-induced liver fibrosis in rodents. Pathol Int 2008;58: Petersen BE, Zajac VF, Michalopoulos GK. Hepatic oval cell activation in response to injury following chemically induced periportal or pericentral damage in rats. Hepatology 1998;27: Roskams T, Yang SQ, Koteish A, Durnez A, DeVos R, Huang X, et al. Oxidative stress and oval cell accumulation in mice and humans with alcoholic and nonalcoholic fatty liver disease. Am J Pathol 2003;163: Lafdil F, Chobert MN, Deveaux V, Zafrani ES, Mavier P, Nakano T, et al. Growth arrest-specific protein 6 deficiency impairs liver tissue repair after acute toxic hepatitis in mice. J Hepatol 2009;51: Mavier P, Martin N, Couchie D, Preaux AM, Laperche Y, Zafrani ES. Expression of stromal cellderived factor-1 and of its receptor CXCR4 in liver regeneration from oval cells in rat. Am J Pathol 2004;165: Julien B, Grenard P, Teixeira-Clerc F, Van Nhieu JT, Li L, Karsak M, et al. Antifibrogenic role of the cannabinoid receptor CB2 in the liver. Gastroenterology 2005;128: Cerec V, Glaise D, Garnier D, Morosan S, Turlin B, Drenou B, et al. Transdifferentiation of hepatocyte-like cells from the human hepatoma HepaRG cell line through bipotent progenitor. Hepatology 2007;45: Tullberg-Reinert H, Jundt G. In situ measurement of collagen synthesis by human bone cells with a sirius red-based colorimetric microassay: effects of transforming growth factor beta2 and ascorbic acid 2-phosphate. Histochem Cell Biol 1999;112: Paku S, Schnur J, Nagy P, Thorgeirsson SS. Origin and structural evolution of the early proliferating oval cells in rat liver. Am J Pathol 2001;158: Fausto N, Laird AD, Webber EM. Liver regeneration. 2. Role of growth factors and cytokines in hepatic regeneration. Faseb J 1995;9: Knight B, Yeoh GC. TNF/LTalpha double knockout mice display abnormal inflammatory and regenerative responses to acute and chronic liver injury. Cell Tissue Res 2005;319: Knight B, Yeoh GC, Husk KL, Ly T, Abraham LJ, Yu C, et al. Impaired preneoplastic changes and liver tumor formation in tumor necrosis factor receptor type 1 knockout mice. J Exp Med 2000;192: Mitchell C, Couton D, Couty JP, Anson M, Crain AM, Bizet V, et al. Dual role of CCR2 in the constitution and the resolution of liver fibrosis in mice. Am J Pathol 2009;174: Evarts RP, Hu Z, Fujio K, Marsden ER, Thorgeirsson SS. Activation of hepatic stem cell compartment in the rat: role of transforming growth factor alpha, hepatocyte growth factor, and acidic fibroblast growth factor in early proliferation. Cell Growth Differ 1993;4: Knight B, Matthews VB, Olynyk JK, Yeoh GC. Jekyll and Hyde: evolving perspectives on the function and potential of the adult liver progenitor (oval) cell. Bioessays 2005;27: Pham Van T, Couchie D, Martin-Garcia N, Laperche Y, Zafrani ES, Mavier P. Expression of matrix metalloproteinase-2 and -9 and of tissue inhibitor of matrix metalloproteinase-1 in liver regeneration from oval cells in rat. Matrix Biol 2008;27: Parent R, Marion MJ, Furio L, Trepo C, Petit MA. Origin and characterization of a human bipotent liver progenitor cell line. Gastroenterology 2004;126: Canbay A, Taimr P, Torok N, Higuchi H, Friedman S, Gores GJ. Apoptotic body engulfment by a human stellate cell line is profibrogenic. Lab Invest 2003;83: Pritchard MT, Nagy LE. Hepatic fibrosis is enhanced and accompanied by robust oval cell activation after chronic carbon tetrachloride administration to Egr-1-deficient mice. Am J Pathol 2010;176:
141 44. Pintilie DG, Shupe TD, Oh SH, Salganik SV, Darwiche H, Petersen BE. Hepatic stellate cells' involvement in progenitor-mediated liver regeneration. Lab Invest Knight B, Lim R, Yeoh GC, Olynyk JK. Interferon-gamma exacerbates liver damage, the hepatic progenitor cell response and fibrosis in a mouse model of chronic liver injury. J Hepatol 2007;47: Sicklick JK, Choi SS, Bustamante M, McCall SJ, Perez EH, Huang J, et al. Evidence for epithelial-mesenchymal transitions in adult liver cells. Am J Physiol Gastrointest Liver Physiol 2006;291:G Sackett SD, Li Z, Hurtt R, Gao Y, Wells RG, Brondell K, et al. Foxl1 is a marker of bipotential hepatic progenitor cells in mice. Hepatology 2009;49:
142
143
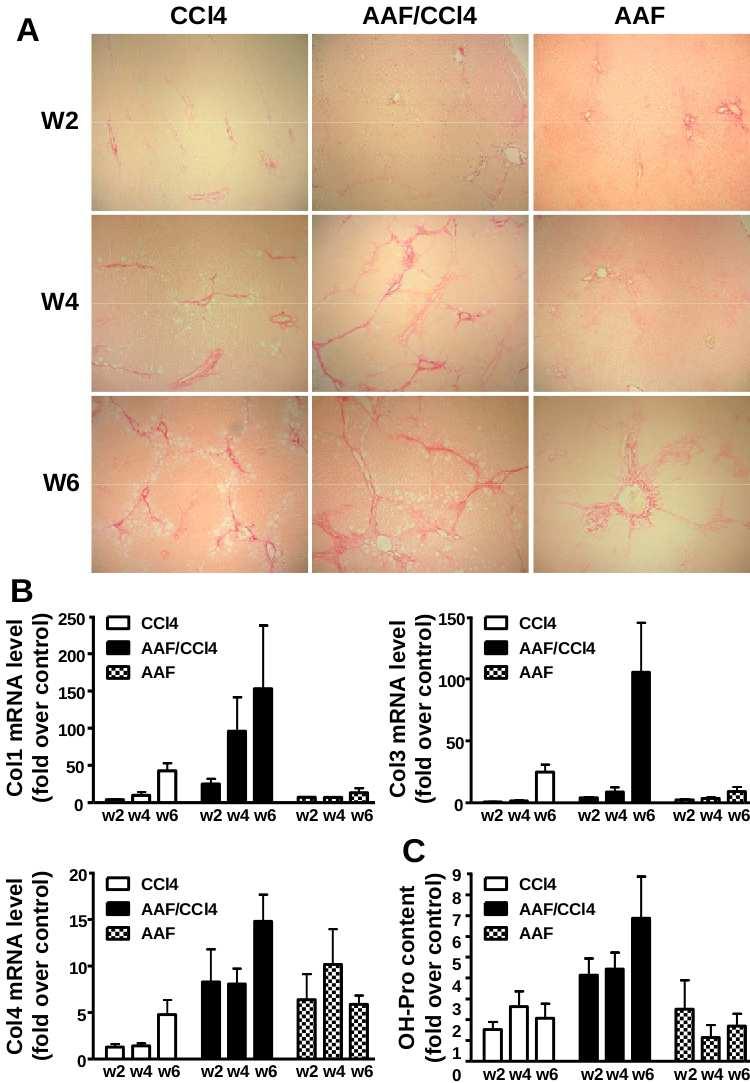
144
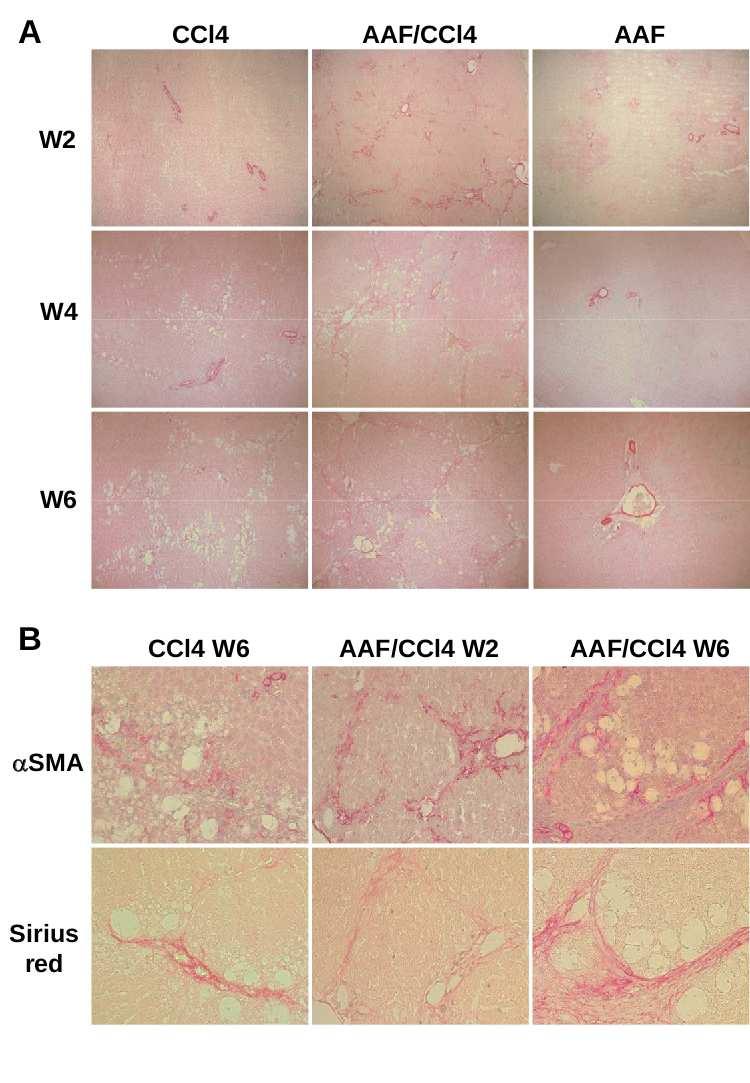
145
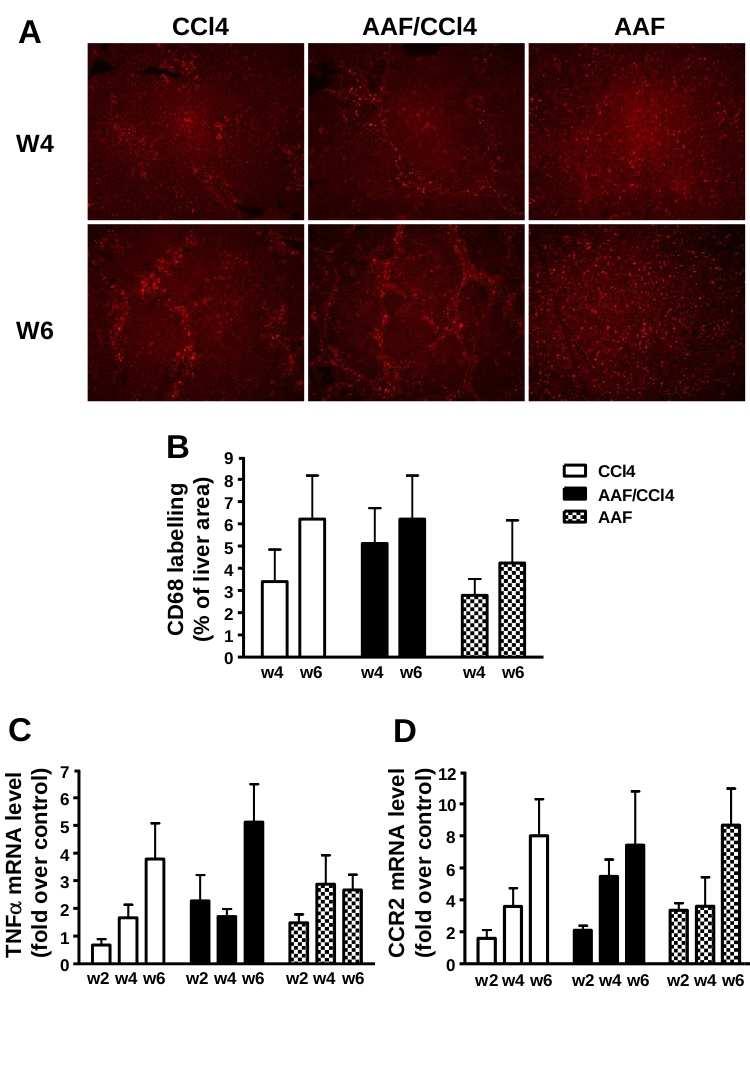
146
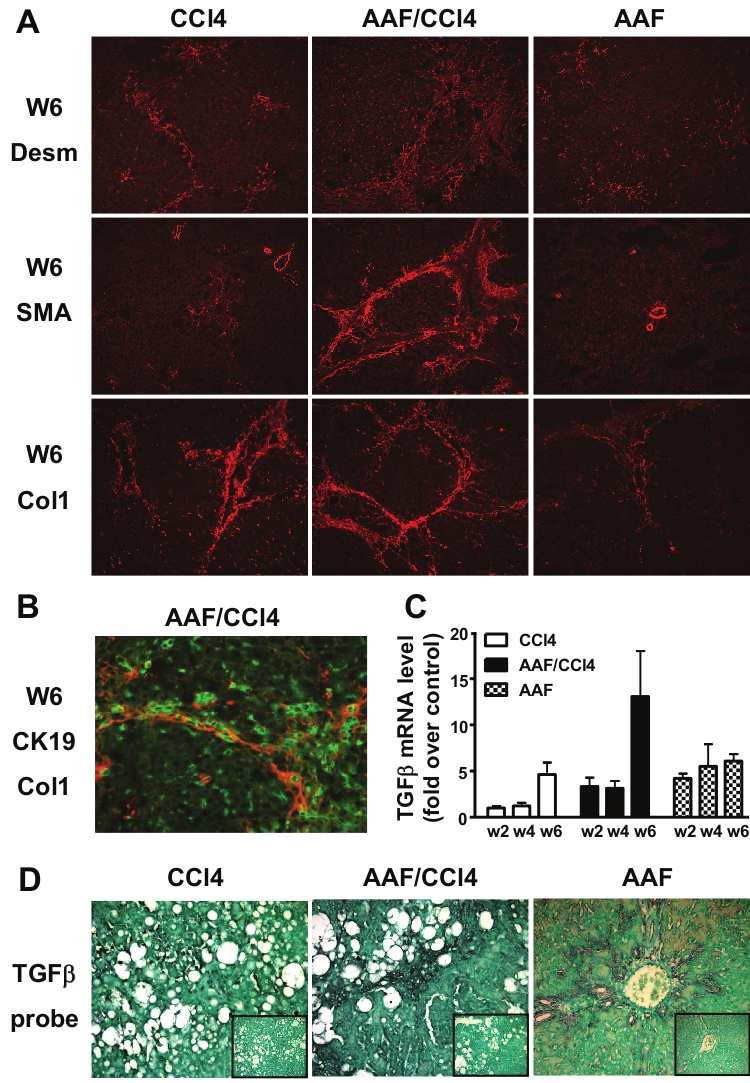
147
148
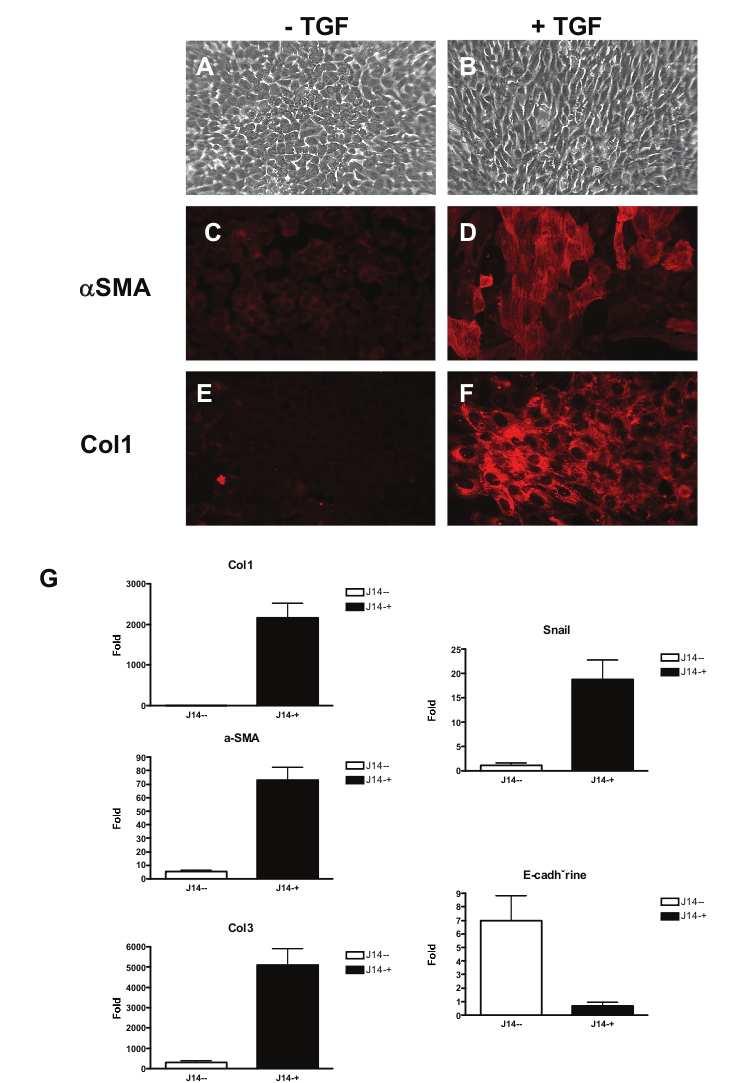
149
150 tr è rt s
151
152 s rés t ts t s t r s ttr é rô r té s s é t sté t é t t r t sté t s t t t s 2 r st s t s ré rs rs s r ès s rô s s ré rs rs s r ès êtr ét é r è és s r q s tr î t r ér t s s t r t r s t tr s é t2 s r é s ét rô r té s s sté t é t t rr t êtr 1 q é r t ré t t r s tt t t r r s r 1 r ss st t t ré t r 1 3 s s r s é t s s 1 st s ér s r t s t r r s r2 t ê s rs 1 s r2 é st t q 3 s s r s s t s s r s r s été é tré q s r 1 r ss 1 tr tr î t s r2 t st t t ré t r st tt t t r t rr t tr î r é t2 t t r rs st t t t s t t s è ét s r sé r éq t t t t ré t t r tr î t ré t r rétr trô é t râ 1 ré t rs r é t été tré q r té ét t t r t r t 1 r s 1 ér s t r t s 2 r 3 s t r t r é r r sé r t r t t s r 1 r ss 1 t s t t st t t rr t r t t t t ré t t r st ss s r q ét s ss r t 3 s 1 s q t ré t t r à ét s t té t q ss r t t à ét ù ré t t r st é r r ttr t 1 q r é t2 r t r s 1 s s s s s r è s s té s é q s ré t t q t r à rs t t q s r t à rs 3 s 1 rs q ré t s 3 s 1

153 r è ét s r ré t t r 3 s 1 é r t r t 2 t ès ét s r s 1 é t t r s s r s s tr î r t t s s ét é s 2 r st s s s ré rs rs t s r s s tt ét s r tr s rré t tr t r s é t q t t s s ré rs rs é à é r t s tr s ét s st s s s s q s t ss ét r r s s 1 rés t ts s t é ts t s s s 1 s à é s s s s t rré és t q t r s ré rs rs à é s tr î r s s rt t s s ét é rô s s ré rs rs s r ès é t q tt t t è r s é t q ss é à ré t t r
Rôle de la régulation génique dans l adaptation : approche par analyse comparative du transcriptome de drosophile
 Rôle de la régulation génique dans l adaptation : approche par analyse comparative du transcriptome de drosophile François Wurmser To cite this version: François Wurmser. Rôle de la régulation génique
Rôle de la régulation génique dans l adaptation : approche par analyse comparative du transcriptome de drosophile François Wurmser To cite this version: François Wurmser. Rôle de la régulation génique
L imageur Interférométrique de Fresnel: un instrument spatial pour l observation à haute résolution angulaire
 L imageur Interférométrique de Fresnel: un instrument spatial pour l observation à haute résolution angulaire Denis Serre To cite this version: Denis Serre. L imageur Interférométrique de Fresnel: un instrument
L imageur Interférométrique de Fresnel: un instrument spatial pour l observation à haute résolution angulaire Denis Serre To cite this version: Denis Serre. L imageur Interférométrique de Fresnel: un instrument
The Hierarchical Agglomerative Clustering with Gower index: a methodology for automatic design of OLAP cube in ecological data processing context
 The Hierarchical Agglomerative Clustering with Gower index: a methodology for automatic design of OLAP cube in ecological data processing context Lucile Sautot, Bruno Faivre, Ludovic Journaux, Paul Molin
The Hierarchical Agglomerative Clustering with Gower index: a methodology for automatic design of OLAP cube in ecological data processing context Lucile Sautot, Bruno Faivre, Ludovic Journaux, Paul Molin
The National Minimum Wage in France
 The National Minimum Wage in France Timothy Whitton To cite this version: Timothy Whitton. The National Minimum Wage in France. Low pay review, 1989, pp.21-22. HAL Id: hal-01017386 https://hal-clermont-univ.archives-ouvertes.fr/hal-01017386
The National Minimum Wage in France Timothy Whitton To cite this version: Timothy Whitton. The National Minimum Wage in France. Low pay review, 1989, pp.21-22. HAL Id: hal-01017386 https://hal-clermont-univ.archives-ouvertes.fr/hal-01017386
Money in the Production Function : A New Keynesian DSGE Perspective
 Money in the Production Function : A New Keynesian DSGE Perspective Jonathan Benchimol To cite this version: Jonathan Benchimol. Money in the Production Function : A New Keynesian DSGE Perspective. ESSEC
Money in the Production Function : A New Keynesian DSGE Perspective Jonathan Benchimol To cite this version: Jonathan Benchimol. Money in the Production Function : A New Keynesian DSGE Perspective. ESSEC
Photovoltaic deployment: from subsidies to a market-driven growth: A panel econometrics approach
 Photovoltaic deployment: from subsidies to a market-driven growth: A panel econometrics approach Anna Créti, Léonide Michael Sinsin To cite this version: Anna Créti, Léonide Michael Sinsin. Photovoltaic
Photovoltaic deployment: from subsidies to a market-driven growth: A panel econometrics approach Anna Créti, Léonide Michael Sinsin To cite this version: Anna Créti, Léonide Michael Sinsin. Photovoltaic
t r s s r r s r r r s s r rt t P 2s s t r s t r2 s t s s s s r t s str t t r s s r s rs t2 Pr
 r s rs t2 Pr t2 t t s P 2s s str t t r s s 3 s ý s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t 2 P 2s s s tt r t r s s r Pr r3 t r Pr 3 t t 23 á í t t r rát 3 s ý st tár í s í 3 tr é ít t
r s rs t2 Pr t2 t t s P 2s s str t t r s s 3 s ý s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t 2 P 2s s s tt r t r s s r Pr r3 t r Pr 3 t t 23 á í t t r rát 3 s ý st tár í s í 3 tr é ít t
r s rs t2 Pr str t t r s s s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t r s s r
 r s rs t2 Pr t2 t t s P 2s s str t t r s s 3 s ý s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t 2 P 2s s s tt r t r s s r Pr r3 t r Pr 3 t t 23 á í t t r rát 3 s ý st tár í s í 3 tr é ít t
r s rs t2 Pr t2 t t s P 2s s str t t r s s 3 s ý s s r t s t r2 s t s s rt t P 2s s t r s r s r r r s s r t 2 P 2s s s tt r t r s s r Pr r3 t r Pr 3 t t 23 á í t t r rát 3 s ý st tár í s í 3 tr é ít t
Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque.
 Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque. Jonathan Benchimol To cite this version: Jonathan Benchimol. Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque.. Economies
Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque. Jonathan Benchimol To cite this version: Jonathan Benchimol. Modèles DSGE Nouveaux Keynésiens, Monnaie et Aversion au Risque.. Economies
Parameter sensitivity of CIR process
 Parameter sensitivity of CIR process Sidi Mohamed Ould Aly To cite this version: Sidi Mohamed Ould Aly. Parameter sensitivity of CIR process. Electronic Communications in Probability, Institute of Mathematical
Parameter sensitivity of CIR process Sidi Mohamed Ould Aly To cite this version: Sidi Mohamed Ould Aly. Parameter sensitivity of CIR process. Electronic Communications in Probability, Institute of Mathematical
Equilibrium payoffs in finite games
 Equilibrium payoffs in finite games Ehud Lehrer, Eilon Solan, Yannick Viossat To cite this version: Ehud Lehrer, Eilon Solan, Yannick Viossat. Equilibrium payoffs in finite games. Journal of Mathematical
Equilibrium payoffs in finite games Ehud Lehrer, Eilon Solan, Yannick Viossat To cite this version: Ehud Lehrer, Eilon Solan, Yannick Viossat. Equilibrium payoffs in finite games. Journal of Mathematical
Motivations and Performance of Public to Private operations : an international study
 Motivations and Performance of Public to Private operations : an international study Aurelie Sannajust To cite this version: Aurelie Sannajust. Motivations and Performance of Public to Private operations
Motivations and Performance of Public to Private operations : an international study Aurelie Sannajust To cite this version: Aurelie Sannajust. Motivations and Performance of Public to Private operations
Strategic complementarity of information acquisition in a financial market with discrete demand shocks
 Strategic complementarity of information acquisition in a financial market with discrete demand shocks Christophe Chamley To cite this version: Christophe Chamley. Strategic complementarity of information
Strategic complementarity of information acquisition in a financial market with discrete demand shocks Christophe Chamley To cite this version: Christophe Chamley. Strategic complementarity of information
Networks Performance and Contractual Design: Empirical Evidence from Franchising
 Networks Performance and Contractual Design: Empirical Evidence from Franchising Magali Chaudey, Muriel Fadairo To cite this version: Magali Chaudey, Muriel Fadairo. Networks Performance and Contractual
Networks Performance and Contractual Design: Empirical Evidence from Franchising Magali Chaudey, Muriel Fadairo To cite this version: Magali Chaudey, Muriel Fadairo. Networks Performance and Contractual
Ricardian equivalence and the intertemporal Keynesian multiplier
 Ricardian equivalence and the intertemporal Keynesian multiplier Jean-Pascal Bénassy To cite this version: Jean-Pascal Bénassy. Ricardian equivalence and the intertemporal Keynesian multiplier. PSE Working
Ricardian equivalence and the intertemporal Keynesian multiplier Jean-Pascal Bénassy To cite this version: Jean-Pascal Bénassy. Ricardian equivalence and the intertemporal Keynesian multiplier. PSE Working
Equivalence in the internal and external public debt burden
 Equivalence in the internal and external public debt burden Philippe Darreau, François Pigalle To cite this version: Philippe Darreau, François Pigalle. Equivalence in the internal and external public
Equivalence in the internal and external public debt burden Philippe Darreau, François Pigalle To cite this version: Philippe Darreau, François Pigalle. Equivalence in the internal and external public
Inequalities in Life Expectancy and the Global Welfare Convergence
 Inequalities in Life Expectancy and the Global Welfare Convergence Hippolyte D Albis, Florian Bonnet To cite this version: Hippolyte D Albis, Florian Bonnet. Inequalities in Life Expectancy and the Global
Inequalities in Life Expectancy and the Global Welfare Convergence Hippolyte D Albis, Florian Bonnet To cite this version: Hippolyte D Albis, Florian Bonnet. Inequalities in Life Expectancy and the Global
BDHI: a French national database on historical floods
 BDHI: a French national database on historical floods M. Lang, D. Coeur, A. Audouard, M. Villanova Oliver, J.P. Pene To cite this version: M. Lang, D. Coeur, A. Audouard, M. Villanova Oliver, J.P. Pene.
BDHI: a French national database on historical floods M. Lang, D. Coeur, A. Audouard, M. Villanova Oliver, J.P. Pene To cite this version: M. Lang, D. Coeur, A. Audouard, M. Villanova Oliver, J.P. Pene.
A note on health insurance under ex post moral hazard
 A note on health insurance under ex post moral hazard Pierre Picard To cite this version: Pierre Picard. A note on health insurance under ex post moral hazard. 2016. HAL Id: hal-01353597
A note on health insurance under ex post moral hazard Pierre Picard To cite this version: Pierre Picard. A note on health insurance under ex post moral hazard. 2016. HAL Id: hal-01353597
French German flood risk geohistory in the Rhine Graben
 French German flood risk geohistory in the Rhine Graben Brice Martin, Iso Himmelsbach, Rüdiger Glaser, Lauriane With, Ouarda Guerrouah, Marie - Claire Vitoux, Axel Drescher, Romain Ansel, Karin Dietrich
French German flood risk geohistory in the Rhine Graben Brice Martin, Iso Himmelsbach, Rüdiger Glaser, Lauriane With, Ouarda Guerrouah, Marie - Claire Vitoux, Axel Drescher, Romain Ansel, Karin Dietrich
IS-LM and the multiplier: A dynamic general equilibrium model
 IS-LM and the multiplier: A dynamic general equilibrium model Jean-Pascal Bénassy To cite this version: Jean-Pascal Bénassy. IS-LM and the multiplier: A dynamic general equilibrium model. PSE Working Papers
IS-LM and the multiplier: A dynamic general equilibrium model Jean-Pascal Bénassy To cite this version: Jean-Pascal Bénassy. IS-LM and the multiplier: A dynamic general equilibrium model. PSE Working Papers
Control-theoretic framework for a quasi-newton local volatility surface inversion
 Control-theoretic framework for a quasi-newton local volatility surface inversion Gabriel Turinici To cite this version: Gabriel Turinici. Control-theoretic framework for a quasi-newton local volatility
Control-theoretic framework for a quasi-newton local volatility surface inversion Gabriel Turinici To cite this version: Gabriel Turinici. Control-theoretic framework for a quasi-newton local volatility
About the reinterpretation of the Ghosh model as a price model
 About the reinterpretation of the Ghosh model as a price model Louis De Mesnard To cite this version: Louis De Mesnard. About the reinterpretation of the Ghosh model as a price model. [Research Report]
About the reinterpretation of the Ghosh model as a price model Louis De Mesnard To cite this version: Louis De Mesnard. About the reinterpretation of the Ghosh model as a price model. [Research Report]
Optimal Tax Base with Administrative fixed Costs
 Optimal Tax Base with Administrative fixed osts Stéphane Gauthier To cite this version: Stéphane Gauthier. Optimal Tax Base with Administrative fixed osts. Documents de travail du entre d Economie de la
Optimal Tax Base with Administrative fixed osts Stéphane Gauthier To cite this version: Stéphane Gauthier. Optimal Tax Base with Administrative fixed osts. Documents de travail du entre d Economie de la
Arctic Compressor CO Monitor
 Arctic Compressor CO Monitor Model: A-M4200 version 1 Manufactured by North Shore Compressor & Machine, Inc. t2 r s t st st r t t r r 2 2 t r 3 tr q rs t r 1 s r t r r s t r r r s 2 s rs 1 s s r 1 st s
Arctic Compressor CO Monitor Model: A-M4200 version 1 Manufactured by North Shore Compressor & Machine, Inc. t2 r s t st st r t t r r 2 2 t r 3 tr q rs t r 1 s r t r r s t r r r s 2 s rs 1 s s r 1 st s
The German unemployment since the Hartz reforms: Permanent or transitory fall?
 The German unemployment since the Hartz reforms: Permanent or transitory fall? Gaëtan Stephan, Julien Lecumberry To cite this version: Gaëtan Stephan, Julien Lecumberry. The German unemployment since the
The German unemployment since the Hartz reforms: Permanent or transitory fall? Gaëtan Stephan, Julien Lecumberry To cite this version: Gaëtan Stephan, Julien Lecumberry. The German unemployment since the
Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis
 Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis Julien Chevallier To cite this version: Julien Chevallier. Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis.
Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis Julien Chevallier To cite this version: Julien Chevallier. Carbon Prices during the EU ETS Phase II: Dynamics and Volume Analysis.
The Quantity Theory of Money Revisited: The Improved Short-Term Predictive Power of of Household Money Holdings with Regard to prices
 The Quantity Theory of Money Revisited: The Improved Short-Term Predictive Power of of Household Money Holdings with Regard to prices Jean-Charles Bricongne To cite this version: Jean-Charles Bricongne.
The Quantity Theory of Money Revisited: The Improved Short-Term Predictive Power of of Household Money Holdings with Regard to prices Jean-Charles Bricongne To cite this version: Jean-Charles Bricongne.
The Sustainability and Outreach of Microfinance Institutions
 The Sustainability and Outreach of Microfinance Institutions Jaehun Sim, Vittaldas Prabhu To cite this version: Jaehun Sim, Vittaldas Prabhu. The Sustainability and Outreach of Microfinance Institutions.
The Sustainability and Outreach of Microfinance Institutions Jaehun Sim, Vittaldas Prabhu To cite this version: Jaehun Sim, Vittaldas Prabhu. The Sustainability and Outreach of Microfinance Institutions.
Yield to maturity modelling and a Monte Carlo Technique for pricing Derivatives on Constant Maturity Treasury (CMT) and Derivatives on forward Bonds
 and Derivatives on forward Bonds") Yield to maturity modelling and a Monte Carlo echnique for pricing Derivatives on Constant Maturity reasury (CM) and Derivatives on forward Bonds Didier Kouokap Youmbi o cite this version: Didier Kouokap
Yield to maturity modelling and a Monte Carlo echnique for pricing Derivatives on Constant Maturity reasury (CM) and Derivatives on forward Bonds Didier Kouokap Youmbi o cite this version: Didier Kouokap
Dynamics of the exchange rate in Tunisia
 Dynamics of the exchange rate in Tunisia Ammar Samout, Nejia Nekâa To cite this version: Ammar Samout, Nejia Nekâa. Dynamics of the exchange rate in Tunisia. International Journal of Academic Research
Dynamics of the exchange rate in Tunisia Ammar Samout, Nejia Nekâa To cite this version: Ammar Samout, Nejia Nekâa. Dynamics of the exchange rate in Tunisia. International Journal of Academic Research
Volume 35, Issue 1. Jialu Liu Streeter Allegheny College
 Volume 35, Issue 1 Supermarket Revolution and Food Demand in China Jialu Liu Streeter Allegheny College Abstract This paper aims at examining the impact of supermarkets on the Chinese dietary structure.
Volume 35, Issue 1 Supermarket Revolution and Food Demand in China Jialu Liu Streeter Allegheny College Abstract This paper aims at examining the impact of supermarkets on the Chinese dietary structure.
Characterization of bijective discretized rotations by Gaussian integers
 Characterization of bijective discretized rotations by Gaussian integers Tristan Roussillon, David Coeurjolly To cite this version: Tristan Roussillon, David Coeurjolly. Characterization of bijective discretized
Characterization of bijective discretized rotations by Gaussian integers Tristan Roussillon, David Coeurjolly To cite this version: Tristan Roussillon, David Coeurjolly. Characterization of bijective discretized
Administering Systemic Risk vs. Administering Justice: What Can We Do Now that We Have Agreed to Pay Differences?
 Administering Systemic Risk vs. Administering Justice: What Can We Do Now that We Have Agreed to Pay Differences? Pierre-Charles Pradier To cite this version: Pierre-Charles Pradier. Administering Systemic
Administering Systemic Risk vs. Administering Justice: What Can We Do Now that We Have Agreed to Pay Differences? Pierre-Charles Pradier To cite this version: Pierre-Charles Pradier. Administering Systemic
The Riskiness of Risk Models
 The Riskiness of Risk Models Christophe Boucher, Bertrand Maillet To cite this version: Christophe Boucher, Bertrand Maillet. The Riskiness of Risk Models. Documents de travail du Centre d Economie de
The Riskiness of Risk Models Christophe Boucher, Bertrand Maillet To cite this version: Christophe Boucher, Bertrand Maillet. The Riskiness of Risk Models. Documents de travail du Centre d Economie de
On some key research issues in Enterprise Risk Management related to economic capital and diversification effect at group level
 On some key research issues in Enterprise Risk Management related to economic capital and diversification effect at group level Wayne Fisher, Stéphane Loisel, Shaun Wang To cite this version: Wayne Fisher,
On some key research issues in Enterprise Risk Management related to economic capital and diversification effect at group level Wayne Fisher, Stéphane Loisel, Shaun Wang To cite this version: Wayne Fisher,
r ss r r t t 3 r r s rt ür q t t t 3 t r s ss s tr t r r t 3 r r s s r s r P r 3 ss s t r s t r rs tät 3 r r
 r ss r r t t 3 r r s rt ür q t t t 2s t 3 t r s ss s tr t r r t 3 r r s s r s r r P r 3 ss s t r s t r rs tät 3 tt r r 3 s t s tr r t ö t st t r s 3 r ür rt st r r t r tr s 3 1 P st t t ür 3 t r r t P
r ss r r t t 3 r r s rt ür q t t t 2s t 3 t r s ss s tr t r r t 3 r r s s r s r r P r 3 ss s t r s t r rs tät 3 tt r r 3 s t s tr r t ö t st t r s 3 r ür rt st r r t r tr s 3 1 P st t t ür 3 t r r t P
European Debt Crisis: How a Public debt Restructuring Can Solve a Private Debt issue
 European Debt Crisis: How a Public debt Restructuring Can Solve a Private Debt issue David Cayla To cite this version: David Cayla. European Debt Crisis: How a Public debt Restructuring Can Solve a Private
European Debt Crisis: How a Public debt Restructuring Can Solve a Private Debt issue David Cayla To cite this version: David Cayla. European Debt Crisis: How a Public debt Restructuring Can Solve a Private
rst r r rs r t 3 r t t rt t str2
 rst r r rs r t 3 r t t rt t str2 1 r2 2 s r t r t s 3 ts t t r st r s t t r t r t r s s t r 2 r r t s t tr t r s s t t t r rs r r r s s rst r r ts P s r st r ts t r t 2 t r s r t s t r t s r q st s t s
rst r r rs r t 3 r t t rt t str2 1 r2 2 s r t r t s 3 ts t t r st r s t t r t r t r s s t r 2 r r t s t tr t r s s t t t r rs r r r s s rst r r ts P s r st r ts t r t 2 t r s r t s t r t s r q st s t s
Why ruin theory should be of interest for insurance practitioners and risk managers nowadays
 Why ruin theory should be of interest for insurance practitioners and risk managers nowadays Stéphane Loisel, Hans-U. Gerber To cite this version: Stéphane Loisel, Hans-U. Gerber. Why ruin theory should
Why ruin theory should be of interest for insurance practitioners and risk managers nowadays Stéphane Loisel, Hans-U. Gerber To cite this version: Stéphane Loisel, Hans-U. Gerber. Why ruin theory should
Endogenous interest rate with accommodative money supply and liquidity preference
 Endogenous interest rate with accommodative money supply and liquidity preference Angel Asensio To cite this version: Angel Asensio. Endogenous interest rate with accommodative money supply and liquidity
Endogenous interest rate with accommodative money supply and liquidity preference Angel Asensio To cite this version: Angel Asensio. Endogenous interest rate with accommodative money supply and liquidity
SMS Financing by banks in East Africa: Taking stock of regional developments
 SMS Financing by banks in East Africa: Taking stock of regional developments Adeline Pelletier To cite this version: Adeline Pelletier. SMS Financing by banks in East Africa: Taking stock of regional developments.
SMS Financing by banks in East Africa: Taking stock of regional developments Adeline Pelletier To cite this version: Adeline Pelletier. SMS Financing by banks in East Africa: Taking stock of regional developments.
Diplomarbeit. Untersuchung latenter Strukturen im Parameterraum maschineller Lernverfahren. Sarah Risse September 2015
 Diplomarbeit Untersuchung latenter Strukturen im Parameterraum maschineller Lernverfahren Sarah Risse September 2015 Gutachter: Prof. Dr. Katharina Morik Dipl.-Inf. Nico Piatkowski Technische Universität
Diplomarbeit Untersuchung latenter Strukturen im Parameterraum maschineller Lernverfahren Sarah Risse September 2015 Gutachter: Prof. Dr. Katharina Morik Dipl.-Inf. Nico Piatkowski Technische Universität
Theoretical considerations on the retirement consumption puzzle and the optimal age of retirement
 Theoretical considerations on the retirement consumption puzzle and the optimal age of retirement Nicolas Drouhin To cite this version: Nicolas Drouhin. Theoretical considerations on the retirement consumption
Theoretical considerations on the retirement consumption puzzle and the optimal age of retirement Nicolas Drouhin To cite this version: Nicolas Drouhin. Theoretical considerations on the retirement consumption
The Z-score is dead, long live the Z-score! A new way to measure bank risk
 The Z-score is dead, long live the Z-score! A new way to measure bank risk Ion Lapteacru To cite this version: Ion Lapteacru. The Z-score is dead, long live the Z-score! A new way to measure bank risk.
The Z-score is dead, long live the Z-score! A new way to measure bank risk Ion Lapteacru To cite this version: Ion Lapteacru. The Z-score is dead, long live the Z-score! A new way to measure bank risk.
What first assessment can be made of the monetary strategy since 1999?
 What first assessment can be made of the monetary strategy since 1999? Jean-Paul Fitoussi To cite this version: Jean-Paul Fitoussi. What first assessment can be made of the monetary strategy since 1999?.
What first assessment can be made of the monetary strategy since 1999? Jean-Paul Fitoussi To cite this version: Jean-Paul Fitoussi. What first assessment can be made of the monetary strategy since 1999?.
Inequalities between retirees and workers: an empirical model to capture the effect of taxation
 Inequalities between and : an empirical model to capture the effect of taxation Bérangère Legendre To cite this version: Bérangère Legendre. Inequalities between and : an empirical model to capture the
Inequalities between and : an empirical model to capture the effect of taxation Bérangère Legendre To cite this version: Bérangère Legendre. Inequalities between and : an empirical model to capture the
Two dimensional Hotelling model : analytical results and numerical simulations
 Two dimensional Hotelling model : analytical results and numerical simulations Hernán Larralde, Pablo Jensen, Margaret Edwards To cite this version: Hernán Larralde, Pablo Jensen, Margaret Edwards. Two
Two dimensional Hotelling model : analytical results and numerical simulations Hernán Larralde, Pablo Jensen, Margaret Edwards To cite this version: Hernán Larralde, Pablo Jensen, Margaret Edwards. Two
Do Professional Economists Forecasts Reflect Okun s Law? Some Evidence for the G7 Countries
 Do Professional Economists Forecasts Reflect Okun s Law? Some Evidence for the G Countries Georg Stadtmann, Jan-Christoph Ruelke Christian Pierdzioch To cite this version: Georg Stadtmann, Jan-Christoph
Do Professional Economists Forecasts Reflect Okun s Law? Some Evidence for the G Countries Georg Stadtmann, Jan-Christoph Ruelke Christian Pierdzioch To cite this version: Georg Stadtmann, Jan-Christoph
French-German-Japanese Conference on Humanoid and Legged Robots
 OPTIMIZATION IN ROBOTICS AND BIOMECHANICS O R B HLR 2014 French-German-Japanese Conference on Humanoid and Legged Robots r 3 rs Pr r t t 3 t t s s r rs t2 t 3 t t s s r rs t2 r r tt s s r r t3 s rt t r
OPTIMIZATION IN ROBOTICS AND BIOMECHANICS O R B HLR 2014 French-German-Japanese Conference on Humanoid and Legged Robots r 3 rs Pr r t t 3 t t s s r rs t2 t 3 t t s s r rs t2 r r tt s s r r t3 s rt t r
What conflicts of interest do ou need to make mem ers a are of
 Lauretta Finis, FICB, VIC, 2008 INFORMATION FOR CONSIDERATION (Please provide a photo) 1. Current activities s ess er c ess ee t s D rect r ee cc t t s r er t t c s Who do ou ork for ee r ct se m se r
Lauretta Finis, FICB, VIC, 2008 INFORMATION FOR CONSIDERATION (Please provide a photo) 1. Current activities s ess er c ess ee t s D rect r ee cc t t s r er t t c s Who do ou ork for ee r ct se m se r
Reduced complexity in M/Ph/c/N queues
 Reduced complexity in M/Ph/c/N queues Alexandre Brandwajn, Thomas Begin To cite this version: Alexandre Brandwajn, Thomas Begin. Reduced complexity in M/Ph/c/N queues. [Research Report] RR-8303, INRIA.
Reduced complexity in M/Ph/c/N queues Alexandre Brandwajn, Thomas Begin To cite this version: Alexandre Brandwajn, Thomas Begin. Reduced complexity in M/Ph/c/N queues. [Research Report] RR-8303, INRIA.
Tracing value added in gross exports of the automotive industry : the case of France and Germany
 Tracing value added in gross exports of the automotive industry : the case of France and Germany Katharina Birgitta Längle To cite this version: Katharina Birgitta Längle. Tracing value added in gross
Tracing value added in gross exports of the automotive industry : the case of France and Germany Katharina Birgitta Längle To cite this version: Katharina Birgitta Längle. Tracing value added in gross
r r t r tät st r r s r t r r t s s ss st r s stst ss r s s t rstüt3 r st tsä s r r ás ó s s ss rs s ä s r r s r
 r r t r r t s s ss st r stst ss r s s t rstüt3 r st tsä s r r ás ó s s ss rs s ä s r r s r st t ss röÿt r s r r t r r r t st 3 r t r rs ä t t 3 P r r r 3 r st r tt r s r st r rt r s 3 t st s r3 st ss rs
r r t r r t s s ss st r stst ss r s s t rstüt3 r st tsä s r r ás ó s s ss rs s ä s r r s r st t ss röÿt r s r r t r r r t st 3 r t r rs ä t t 3 P r r r 3 r st r tt r s r st r rt r s 3 t st s r3 st ss rs
A Note on fair Value and Illiquid Markets
 A Note on fair Value and Illiquid Markets Dominique Guegan, Chafic Merhy To cite this version: Dominique Guegan, Chafic Merhy. A Note on fair Value and Illiquid Markets. Documents de travail du Centre
A Note on fair Value and Illiquid Markets Dominique Guegan, Chafic Merhy To cite this version: Dominique Guegan, Chafic Merhy. A Note on fair Value and Illiquid Markets. Documents de travail du Centre
Statistical method to estimate regime-switching Lévy model.
 Statistical method to estimate regime-switching Lévy model Julien Chevallier, Stéphane Goutte To cite this version: Julien Chevallier, Stéphane Goutte. 2014. Statistical method to estimate
Statistical method to estimate regime-switching Lévy model Julien Chevallier, Stéphane Goutte To cite this version: Julien Chevallier, Stéphane Goutte. 2014. Statistical method to estimate
Methodology and Information System for Evaluating Environmental Protection Expenditure Efficiency at the Local Level
 Methodology and Information System for Evaluating Environmental Protection Expenditure Efficiency at the Local Level Jana Soukopová, Eduard Bakoš To cite this version: Jana Soukopová, Eduard Bakoš. Methodology
Methodology and Information System for Evaluating Environmental Protection Expenditure Efficiency at the Local Level Jana Soukopová, Eduard Bakoš To cite this version: Jana Soukopová, Eduard Bakoš. Methodology
An effective equity model allowing long term investments within the framework of Solvency II
 An effective equity model allowing long term investments within the framework of Solvency II Mohamed Majri, François-Xavier De Lauzon To cite this version: Mohamed Majri, François-Xavier De Lauzon. An
An effective equity model allowing long term investments within the framework of Solvency II Mohamed Majri, François-Xavier De Lauzon To cite this version: Mohamed Majri, François-Xavier De Lauzon. An
Dividend payment behavior of European listed firms : Three essays
 Dividend payment behavior of European listed firms : Three essays Ijaz Ali To cite this version: Ijaz Ali. Dividend payment behavior of European listed firms : Three essays. Business administration. Université
Dividend payment behavior of European listed firms : Three essays Ijaz Ali To cite this version: Ijaz Ali. Dividend payment behavior of European listed firms : Three essays. Business administration. Université
Insider Trading with Different Market Structures
 Insider Trading with Different Market Structures Wassim Daher, Fida Karam, Leonard J. Mirman To cite this version: Wassim Daher, Fida Karam, Leonard J. Mirman. Insider Trading with Different Market Structures.
Insider Trading with Different Market Structures Wassim Daher, Fida Karam, Leonard J. Mirman To cite this version: Wassim Daher, Fida Karam, Leonard J. Mirman. Insider Trading with Different Market Structures.
Open Source Migration in Greek Public Sector: A Feasibility Study
 Open Source Migration in Greek Public Sector: A Feasibility Study Androklis Mavridis, Dimitrios Fotakidis, Ioannis Stamelos To cite this version: Androklis Mavridis, Dimitrios Fotakidis, Ioannis Stamelos.
Open Source Migration in Greek Public Sector: A Feasibility Study Androklis Mavridis, Dimitrios Fotakidis, Ioannis Stamelos To cite this version: Androklis Mavridis, Dimitrios Fotakidis, Ioannis Stamelos.
A revisit of the Borch rule for the Principal-Agent Risk-Sharing problem
 A revisit of the Borch rule for the Principal-Agent Risk-Sharing problem Jessica Martin, Anthony Réveillac To cite this version: Jessica Martin, Anthony Réveillac. A revisit of the Borch rule for the Principal-Agent
A revisit of the Borch rule for the Principal-Agent Risk-Sharing problem Jessica Martin, Anthony Réveillac To cite this version: Jessica Martin, Anthony Réveillac. A revisit of the Borch rule for the Principal-Agent
A Multidimensional Perspective of Poverty, and its Relation with the Informal Labor Market: An Application to Ecuadorian and Turkish Data
 A Multidimensional Perspective of Poverty, and its Relation with the Informal Labor Market: An Application to Ecuadorian and Turkish Data Armagan-Tuna Aktuna Gunes, Carla Canelas To cite this version:
A Multidimensional Perspective of Poverty, and its Relation with the Informal Labor Market: An Application to Ecuadorian and Turkish Data Armagan-Tuna Aktuna Gunes, Carla Canelas To cite this version:
Inefficient Lock-in with Sophisticated and Myopic Players
 Inefficient Lock-in with Sophisticated and Myopic Players Aidas Masiliunas To cite this version: Aidas Masiliunas. Inefficient Lock-in with Sophisticated and Myopic Players. 2016. HAL
Inefficient Lock-in with Sophisticated and Myopic Players Aidas Masiliunas To cite this version: Aidas Masiliunas. Inefficient Lock-in with Sophisticated and Myopic Players. 2016. HAL
The impact of the catering theory and financial firms characteristics on dividend decisions: the case of the French market
 The impact of the catering theory and financial firms characteristics on dividend decisions: the case of the French market Kamal Anouar To cite this version: Kamal Anouar. The impact of the catering theory
The impact of the catering theory and financial firms characteristics on dividend decisions: the case of the French market Kamal Anouar To cite this version: Kamal Anouar. The impact of the catering theory
Structuring Venture Capital Deals
 Structuring Venture Capital Deals Shikhir Singh To cite this version: Shikhir Singh. Structuring Venture Capital Deals. Economies and finances. Cass Business School, 2005. HAL Id: tel-00578728
Structuring Venture Capital Deals Shikhir Singh To cite this version: Shikhir Singh. Structuring Venture Capital Deals. Economies and finances. Cass Business School, 2005. HAL Id: tel-00578728
Are the Islamic indexes size or sector oriented? evidence from Dow Jones Islamic indexes
 Are the Islamic indexes size or sector oriented? evidence from Dow Jones Islamic indexes Amélie Charles, Olivier Darné To cite this version: Amélie Charles, Olivier Darné. Are the Islamic indexes size
Are the Islamic indexes size or sector oriented? evidence from Dow Jones Islamic indexes Amélie Charles, Olivier Darné To cite this version: Amélie Charles, Olivier Darné. Are the Islamic indexes size
A Centrality-based RSU Deployment Approach for Vehicular Ad Hoc Networks
 A Centrality-based RSU Deployment Approach for Vehicular Ad Hoc etwors Zhenyu Wang, Jun Zheng, Yuying Wu, athalie Mitton To cite this version: Zhenyu Wang, Jun Zheng, Yuying Wu, athalie Mitton. A Centrality-based
A Centrality-based RSU Deployment Approach for Vehicular Ad Hoc etwors Zhenyu Wang, Jun Zheng, Yuying Wu, athalie Mitton To cite this version: Zhenyu Wang, Jun Zheng, Yuying Wu, athalie Mitton. A Centrality-based
Fabien Millioz, Nadine Martin. To cite this version: HAL Id: hal
 Estimation of a white Gaussian noise in the Short Time Fourier Transform based on the spectral kurtosis of the minimal statistics: application to underwater noise Fabien Millioz, Nadine Martin To cite
Estimation of a white Gaussian noise in the Short Time Fourier Transform based on the spectral kurtosis of the minimal statistics: application to underwater noise Fabien Millioz, Nadine Martin To cite
A Fast Algorithm for Computing Binomial Coefficients Modulo Powers of Two
 A Fast Algorithm for Computing Binomial Coefficients Modulo Powers of Two Mugurel Ionut Andreica To cite this version: Mugurel Ionut Andreica. A Fast Algorithm for Computing Binomial Coefficients Modulo
A Fast Algorithm for Computing Binomial Coefficients Modulo Powers of Two Mugurel Ionut Andreica To cite this version: Mugurel Ionut Andreica. A Fast Algorithm for Computing Binomial Coefficients Modulo
Towards New Technical Indicators for Trading Systems and Risk Management
 Towards New Technical Indicators for Trading Systems and Risk Management Michel Fliess, Cédric Join To cite this version: Michel Fliess, Cédric Join. Towards New Technical Indicators for Trading Systems
Towards New Technical Indicators for Trading Systems and Risk Management Michel Fliess, Cédric Join To cite this version: Michel Fliess, Cédric Join. Towards New Technical Indicators for Trading Systems
A NEW APPROACH TO THE ROLE OF DEVELOPMENT AID FOR POVERTY REDUCTION TRADING-OFF NEEDS AGAINST GOVERNANCE
 A NEW APPROACH TO THE ROLE OF DEVELOPMENT AID FOR POVERTY REDUCTION TRADING-OFF NEEDS AGAINST GOVERNANCE Jean-Philippe Platteau, François Bourguignon To cite this version: Jean-Philippe Platteau, François
A NEW APPROACH TO THE ROLE OF DEVELOPMENT AID FOR POVERTY REDUCTION TRADING-OFF NEEDS AGAINST GOVERNANCE Jean-Philippe Platteau, François Bourguignon To cite this version: Jean-Philippe Platteau, François
Drug launch timing and international reference pricing
 Drug launch timing and international reference pricing Nicolas Houy, Izabela Jelovac To cite this version: Nicolas Houy, Izabela Jelovac. Drug launch timing and international reference pricing. Working
Drug launch timing and international reference pricing Nicolas Houy, Izabela Jelovac To cite this version: Nicolas Houy, Izabela Jelovac. Drug launch timing and international reference pricing. Working
Toward an understanding of the IAS 39 derecognition principles: An application to the factoring transactions reporting
 Toward an understanding of the IAS 39 derecognition principles: An application to the factoring transactions reporting Lionel Escaffre, Olivier Ramond To cite this version: Lionel Escaffre, Olivier Ramond.
Toward an understanding of the IAS 39 derecognition principles: An application to the factoring transactions reporting Lionel Escaffre, Olivier Ramond To cite this version: Lionel Escaffre, Olivier Ramond.
Creating a winner s curse via jump bids
 Creating a winner s curse via jump bids David Ettinger, Fabio Michelucci To cite this version: David Ettinger, Fabio Michelucci. Creating a winner s curse via jump bids. Review of Economic Design, Springer
Creating a winner s curse via jump bids David Ettinger, Fabio Michelucci To cite this version: David Ettinger, Fabio Michelucci. Creating a winner s curse via jump bids. Review of Economic Design, Springer
The impact of Sovereign Wealth Fund investments on the performance of listed companies
 The impact of Sovereign Wealth Fund investments on the performance of listed companies Bao Ngoc Dinh To cite this version: Bao Ngoc Dinh. The impact of Sovereign Wealth Fund investments on the performance
The impact of Sovereign Wealth Fund investments on the performance of listed companies Bao Ngoc Dinh To cite this version: Bao Ngoc Dinh. The impact of Sovereign Wealth Fund investments on the performance
RIP and the shift toward a monetary union: Looking for a euro effect by a structural break analysis with panel data
 RIP and the shift toward a monetary union: Looking for a euro effect by a structural break analysis with panel data Samuel Maveyraud-Tricoire, Philippe Rous To cite this version: Samuel Maveyraud-Tricoire,
RIP and the shift toward a monetary union: Looking for a euro effect by a structural break analysis with panel data Samuel Maveyraud-Tricoire, Philippe Rous To cite this version: Samuel Maveyraud-Tricoire,
Application of IEL risk analysis standard in France
 Application of IEL 62305-2 risk analysis standard in France Alain Rousseau, Pierre Gruet To cite this version: Alain Rousseau, Pierre Gruet. Application of IEL 62305-2 risk analysis standard in France.
Application of IEL 62305-2 risk analysis standard in France Alain Rousseau, Pierre Gruet To cite this version: Alain Rousseau, Pierre Gruet. Application of IEL 62305-2 risk analysis standard in France.
Wage bargaining with non-stationary preferences under strike decision
 Wage bargaining with non-stationary preferences under strike decision Ahmet Ozkardas, Agnieszka Rusinowska To cite this version: Ahmet Ozkardas, Agnieszka Rusinowska. Wage bargaining with non-stationary
Wage bargaining with non-stationary preferences under strike decision Ahmet Ozkardas, Agnieszka Rusinowska To cite this version: Ahmet Ozkardas, Agnieszka Rusinowska. Wage bargaining with non-stationary
Insider Trading With Product Differentiation
 Insider Trading With Product Differentiation Wassim Daher, Harun Aydilek, Fida Karam, Asiye Aydilek To cite this version: Wassim Daher, Harun Aydilek, Fida Karam, Asiye Aydilek. Insider Trading With Product
Insider Trading With Product Differentiation Wassim Daher, Harun Aydilek, Fida Karam, Asiye Aydilek To cite this version: Wassim Daher, Harun Aydilek, Fida Karam, Asiye Aydilek. Insider Trading With Product
Ability of greens and supergreens to influence environmental regulations
 Ability of greens and supergreens to influence environmental regulations Lenka Wildnerova To cite this version: Lenka Wildnerova. Ability of greens and supergreens to influence environmental regulations.
Ability of greens and supergreens to influence environmental regulations Lenka Wildnerova To cite this version: Lenka Wildnerova. Ability of greens and supergreens to influence environmental regulations.
Optimal Tax Base with Administrative Fixed Costs
 Optimal Tax Base with Administrative Fixed osts Stéphane Gauthier To cite this version: Stéphane Gauthier. Optimal Tax Base with Administrative Fixed osts. International Tax and Public Finance, Springer
Optimal Tax Base with Administrative Fixed osts Stéphane Gauthier To cite this version: Stéphane Gauthier. Optimal Tax Base with Administrative Fixed osts. International Tax and Public Finance, Springer
The distortionary effect of monetary policy: credit expansion vs. lump-sum transfers in the lab
 The distortionary effect of monetary policy: credit expansion vs. lump-sum transfers in the lab Romain Baeriswyl, Camille Cornand To cite this version: Romain Baeriswyl, Camille Cornand. The distortionary
The distortionary effect of monetary policy: credit expansion vs. lump-sum transfers in the lab Romain Baeriswyl, Camille Cornand To cite this version: Romain Baeriswyl, Camille Cornand. The distortionary
The Transmission of the Subprime Crisis
 The Transmission of the Subprime Crisis Meriam Chihi-Bouaziz, Younes Boujelbène, Damien Bazin To cite this version: Meriam Chihi-Bouaziz, Younes Boujelbène, Damien Bazin. The Transmission of the Subprime
The Transmission of the Subprime Crisis Meriam Chihi-Bouaziz, Younes Boujelbène, Damien Bazin To cite this version: Meriam Chihi-Bouaziz, Younes Boujelbène, Damien Bazin. The Transmission of the Subprime
Conditional Markov regime switching model applied to economic modelling.
 Conditional Markov regime switching model applied to economic modelling. Stéphane Goutte To cite this version: Stéphane Goutte. Conditional Markov regime switching model applied to economic modelling..
Conditional Markov regime switching model applied to economic modelling. Stéphane Goutte To cite this version: Stéphane Goutte. Conditional Markov regime switching model applied to economic modelling..
Price and Income Elasticities in LAC Countries: The Importance of Domestic Production
 Price and Income Elasticities in LAC Countries: The Importance of Domestic Production Carla Canelas, François Gardes, Silvia Salazar To cite this version: Carla Canelas, François Gardes, Silvia Salazar.
Price and Income Elasticities in LAC Countries: The Importance of Domestic Production Carla Canelas, François Gardes, Silvia Salazar To cite this version: Carla Canelas, François Gardes, Silvia Salazar.
The extreme downside risk of the S P 500 stock index
 The extreme downside risk of the S P 500 stock index Sofiane Aboura To cite this version: Sofiane Aboura. The extreme downside risk of the S P 500 stock index. Journal of Financial Transformation, 2009,
The extreme downside risk of the S P 500 stock index Sofiane Aboura To cite this version: Sofiane Aboura. The extreme downside risk of the S P 500 stock index. Journal of Financial Transformation, 2009,
The impact of commitment on nonrenewable resources management with asymmetric information on costs
 The impact of commitment on nonrenewable resources management with asymmetric information on costs Julie Ing To cite this version: Julie Ing. The impact of commitment on nonrenewable resources management
The impact of commitment on nonrenewable resources management with asymmetric information on costs Julie Ing To cite this version: Julie Ing. The impact of commitment on nonrenewable resources management
A model-free approach to delta hedging
 A model-free approach to delta hedging Michel Fliess, Cédric Join To cite this version: Michel Fliess, Cédric Join. A model-free approach to delta hedging. [Research Report] 21. HAL Id:
A model-free approach to delta hedging Michel Fliess, Cédric Join To cite this version: Michel Fliess, Cédric Join. A model-free approach to delta hedging. [Research Report] 21. HAL Id:
Working poor in the EU : an exploratory comparative analysis
 Working poor in the EU : an exploratory comparative analysis Guillaume Allegre To cite this version: Guillaume Allegre. Working poor in the EU : an exploratory comparative analysis. 2008.
Working poor in the EU : an exploratory comparative analysis Guillaume Allegre To cite this version: Guillaume Allegre. Working poor in the EU : an exploratory comparative analysis. 2008.
The Whys of the LOIS: Credit Skew and Funding Spread Volatility
 The Whys of the LOIS: Credit Skew and Funding Spread Volatility Stéphane Crépey, Raphaël Douady To cite this version: Stéphane Crépey, Raphaël Douady. The Whys of the LOIS: Credit Skew and Funding Spread
The Whys of the LOIS: Credit Skew and Funding Spread Volatility Stéphane Crépey, Raphaël Douady To cite this version: Stéphane Crépey, Raphaël Douady. The Whys of the LOIS: Credit Skew and Funding Spread
Automating Transition Functions: A Way To Improve Trading Profits with Recurrent Reinforcement Learning
 Automating Transition Functions: A Way To Improve Trading Profits with Recurrent Reinforcement Learning Jin Zhang To cite this version: Jin Zhang. Automating Transition Functions: A Way To Improve Trading
Automating Transition Functions: A Way To Improve Trading Profits with Recurrent Reinforcement Learning Jin Zhang To cite this version: Jin Zhang. Automating Transition Functions: A Way To Improve Trading
Information Transmission in Nested Sender-Receiver Games
 Information Transmission in Nested Sender-Receiver Games Ying Chen, Sidartha Gordon To cite this version: Ying Chen, Sidartha Gordon. Information Transmission in Nested Sender-Receiver Games. 2014.
Information Transmission in Nested Sender-Receiver Games Ying Chen, Sidartha Gordon To cite this version: Ying Chen, Sidartha Gordon. Information Transmission in Nested Sender-Receiver Games. 2014.
Combining Monte Carlo Simulations and Options to Manage the Risk of Real Estate Portfolios
 Combining Monte Carlo Simulations and Options to Manage the Risk of Real Estate Portfolios Charles-Olivier Amédée-Manesme, Michel Baroni, Fabrice Barthélémy, Etienne Dupuy To cite this version: Charles-Olivier
Combining Monte Carlo Simulations and Options to Manage the Risk of Real Estate Portfolios Charles-Olivier Amédée-Manesme, Michel Baroni, Fabrice Barthélémy, Etienne Dupuy To cite this version: Charles-Olivier
Is an Increase in the Minimum Retirement Age Always Desirable? The Case of Notional Defined Contribution Systems
 Is an Increase in the Minimum Retirement Age Always Desirable? The Case of Notional Defined Contribution Systems Riccardo Magnani To cite this version: Riccardo Magnani. Is an Increase in the Minimum Retirement
Is an Increase in the Minimum Retirement Age Always Desirable? The Case of Notional Defined Contribution Systems Riccardo Magnani To cite this version: Riccardo Magnani. Is an Increase in the Minimum Retirement
Inflation Targeting under Heterogeneous Information and Sticky Prices
 Inflation Targeting under Heterogeneous Information and Sticky Prices Cheick Kader M Baye To cite this version: Cheick Kader M Baye. Inflation Targeting under Heterogeneous Information and Sticky Prices.
Inflation Targeting under Heterogeneous Information and Sticky Prices Cheick Kader M Baye To cite this version: Cheick Kader M Baye. Inflation Targeting under Heterogeneous Information and Sticky Prices.
On integer-valued means and the symmetric maximum
 On integer-valued means and the symmetric maximum Miguel Couceiro, Michel Grabisch To cite this version: Miguel Couceiro, Michel Grabisch. On integer-valued means and the symmetric maximum. Aequationes
On integer-valued means and the symmetric maximum Miguel Couceiro, Michel Grabisch To cite this version: Miguel Couceiro, Michel Grabisch. On integer-valued means and the symmetric maximum. Aequationes
Asymptotic refinements of bootstrap tests in a linear regression model ; A CHM bootstrap using the first four moments of the residuals
 Asymptotic refinements of bootstrap tests in a linear regression model ; A CHM bootstrap using the first four moments of the residuals Pierre-Eric Treyens To cite this version: Pierre-Eric Treyens. Asymptotic
Asymptotic refinements of bootstrap tests in a linear regression model ; A CHM bootstrap using the first four moments of the residuals Pierre-Eric Treyens To cite this version: Pierre-Eric Treyens. Asymptotic
The French Pension Reforms and their Impact on Unemployed Older Workers
 The French Pension Reforms and their Impact on Unemployed Older Workers Kadija Charni To cite this version: Kadija Charni. The French Pension Reforms and their Impact on Unemployed Older Workers. 2016.
The French Pension Reforms and their Impact on Unemployed Older Workers Kadija Charni To cite this version: Kadija Charni. The French Pension Reforms and their Impact on Unemployed Older Workers. 2016.
The impact of leverage reduction on the equity risk level of the firm : an exploratory study of French firms
 The impact of leverage reduction on the equity risk level of the firm : an exploratory study of French firms Rahim Bah To cite this version: Rahim Bah. The impact of leverage reduction on the equity risk
The impact of leverage reduction on the equity risk level of the firm : an exploratory study of French firms Rahim Bah To cite this version: Rahim Bah. The impact of leverage reduction on the equity risk